Основы гистологии. Презентация на тему "основы гистологии". Функции соединительной ткани
 Ткани - это совокупность клеток и неклеточных структур (неклеточных веществ), сходных по происхождению, строению и выполняемым функциям. Выделяют четыре основные группы тканей: эпителиальные, мышечные, соединительные и нервную.
Ткани - это совокупность клеток и неклеточных структур (неклеточных веществ), сходных по происхождению, строению и выполняемым функциям. Выделяют четыре основные группы тканей: эпителиальные, мышечные, соединительные и нервную.




 … Эпителиальные ткани покрывают организм снаружи и выстилают изнутри полые органы и стенки полостей тела. Особый вид эпителиальной ткани - железистый эпителий - образует большинство желез (щитовидную, потовые, печень и др.).
… Эпителиальные ткани покрывают организм снаружи и выстилают изнутри полые органы и стенки полостей тела. Особый вид эпителиальной ткани - железистый эпителий - образует большинство желез (щитовидную, потовые, печень и др.).

 … Эпителиальные ткани имеют следующие особенности: — их клетки тесно прилегают друг к другу, образуя пласт, — межклеточного вещества очень мало; — клетки обладают способностью к восстановлению (регенерации).
… Эпителиальные ткани имеют следующие особенности: — их клетки тесно прилегают друг к другу, образуя пласт, — межклеточного вещества очень мало; — клетки обладают способностью к восстановлению (регенерации).
 … Эпителиальные клетки по форме могут быть плоскими, цилиндрическими, кубическими. По количеству пластов эпителии бывают однослойные и многослойные.
… Эпителиальные клетки по форме могут быть плоскими, цилиндрическими, кубическими. По количеству пластов эпителии бывают однослойные и многослойные.
 … Примеры эпителиев: однослойный плоский выстилает грудную и брюшную полости тела; многослойный плоский образует наружный слой кожи (эпидермис); однослойный цилиндрический выстилает большую часть кишечного тракта; многослойный цилиндрический - полость верхних дыхательных путей); однослойный кубический образует канальцы нефронов почек. Функции эпителиальных тканей; пограничная, защитная, секреторная, всасывания.
… Примеры эпителиев: однослойный плоский выстилает грудную и брюшную полости тела; многослойный плоский образует наружный слой кожи (эпидермис); однослойный цилиндрический выстилает большую часть кишечного тракта; многослойный цилиндрический - полость верхних дыхательных путей); однослойный кубический образует канальцы нефронов почек. Функции эпителиальных тканей; пограничная, защитная, секреторная, всасывания.
 СОЕДИНИТЕЛЬНАЯ ТКАНЬ СОБСТВЕННО СОЕДИНИТЕЛЬНАЯ СКЕЛЕТНАЯ Волокнистая Хрящевая 1. рыхлая 1. гиалиновый хрящ 2. плотная 2. эластический хрящ 3. оформленная 3. волокнистый хрящ 4. неоформленная Со специальными свойствами Костная 1. ретикулярная 1. грубоволокнистая 2. жировая 2. пластинчатая: 3. слизистая компактное вещество 4. пигментная губчатое вещество
СОЕДИНИТЕЛЬНАЯ ТКАНЬ СОБСТВЕННО СОЕДИНИТЕЛЬНАЯ СКЕЛЕТНАЯ Волокнистая Хрящевая 1. рыхлая 1. гиалиновый хрящ 2. плотная 2. эластический хрящ 3. оформленная 3. волокнистый хрящ 4. неоформленная Со специальными свойствами Костная 1. ретикулярная 1. грубоволокнистая 2. жировая 2. пластинчатая: 3. слизистая компактное вещество 4. пигментная губчатое вещество
 … Соединительные ткани (ткани внутренней среды) объединяют группы тканей мезодермального происхождения, очень различных по строению и выполняемым функциям. Виды соединительной ткани: костная, хрящевая, подкожная жировая клетчатка, связки, сухожилия, кровь, лимфа и др.
… Соединительные ткани (ткани внутренней среды) объединяют группы тканей мезодермального происхождения, очень различных по строению и выполняемым функциям. Виды соединительной ткани: костная, хрящевая, подкожная жировая клетчатка, связки, сухожилия, кровь, лимфа и др.


 … Соединительные ткани Общей характерной чертой строения этих тканей является рыхлое расположение клеток, отделенных друг от друга хорошо выраженным межклеточным веществом, которое образовано различными волокнами белковой природы (коллагеновыми, эластическими) и основным аморфным веществом.
… Соединительные ткани Общей характерной чертой строения этих тканей является рыхлое расположение клеток, отделенных друг от друга хорошо выраженным межклеточным веществом, которое образовано различными волокнами белковой природы (коллагеновыми, эластическими) и основным аморфным веществом.
 … Кровь - разновидность соединительной ткани, у которой межклеточное вещество жидкое (плазма), благодаря чему одной из основных функций крови является транспортная (переносит газы, питательные вещества, гормоны, конечные продукты жизнедеятельности клеток и др.).
… Кровь - разновидность соединительной ткани, у которой межклеточное вещество жидкое (плазма), благодаря чему одной из основных функций крови является транспортная (переносит газы, питательные вещества, гормоны, конечные продукты жизнедеятельности клеток и др.).
 … Межклеточное вещество рыхлой волокнистой соединительной ткани, находящейся в прослойках между органами, а также соединяющей кожу с мышцами, состоит из аморфного вещества и свободно расположенных в разных направлениях эластических волокон. Благодаря такому строению межклеточного вещества кожа подвижна. Эта ткань выполняет опорную, защитную и питательную функции.
… Межклеточное вещество рыхлой волокнистой соединительной ткани, находящейся в прослойках между органами, а также соединяющей кожу с мышцами, состоит из аморфного вещества и свободно расположенных в разных направлениях эластических волокон. Благодаря такому строению межклеточного вещества кожа подвижна. Эта ткань выполняет опорную, защитную и питательную функции.



 … Мышечные ткани обусловливают все виды двигательных процессов внутри организма, а также перемещение организма и его частей в пространстве.
… Мышечные ткани обусловливают все виды двигательных процессов внутри организма, а также перемещение организма и его частей в пространстве.
 … Это обеспечивается за счет особых свойств мышечных клеток - возбудимости и сократимости. Во всех клетках мышечных тканей содержатся тончайшие сократительные волоконца - миофибриллы, образованные линейными молекулами белков - актином и миозином. При скольжении их относительно друга происходит изменение длины мышечных клеток.
… Это обеспечивается за счет особых свойств мышечных клеток - возбудимости и сократимости. Во всех клетках мышечных тканей содержатся тончайшие сократительные волоконца - миофибриллы, образованные линейными молекулами белков - актином и миозином. При скольжении их относительно друга происходит изменение длины мышечных клеток.
 … Поперечнополосатая (скелетная) мышечная ткань построена из множества многоядерных волокноподобных клеток длиной 1- 12 см. Из нее построены все скелетные мышцы, мышцы языка, стенок ротовой полости, глотки, гортани, верхней части пищевода, мимические, диафрагма. Рисунок 1. Волокна поперечнополосатой мышечной ткани: а) внешний вид волокон; б) поперечный разрез волокон
… Поперечнополосатая (скелетная) мышечная ткань построена из множества многоядерных волокноподобных клеток длиной 1- 12 см. Из нее построены все скелетные мышцы, мышцы языка, стенок ротовой полости, глотки, гортани, верхней части пищевода, мимические, диафрагма. Рисунок 1. Волокна поперечнополосатой мышечной ткани: а) внешний вид волокон; б) поперечный разрез волокон
 … Особенности поперечнополосатой мышечной ткани: быстрота и произвольность (т. е. зависимость сокращении от воли, желания человека), потребление большого количества энергии и кислорода, быстрая утомляемость. Рисунок 1. Волокна поперечнополосатой мышечной ткани: а) внешний вид волокон; б) поперечный разрез волокон
… Особенности поперечнополосатой мышечной ткани: быстрота и произвольность (т. е. зависимость сокращении от воли, желания человека), потребление большого количества энергии и кислорода, быстрая утомляемость. Рисунок 1. Волокна поперечнополосатой мышечной ткани: а) внешний вид волокон; б) поперечный разрез волокон
 … Сердечная ткань состоит из поперечно исчерченных одноядерных мышечных клеток, но обладает иными свойствами. Клетки расположены не параллельным пучком, как скелетные, а ветвятся, образуя единую сеть. Благодаря множеству клеточных контактов поступающий нервный импульс передается от одной клетки к другой, обеспечивая одновременное сокращение, а затем расслабление сердечной мышцы, что позволяет ей выполнять насосную функцию.
… Сердечная ткань состоит из поперечно исчерченных одноядерных мышечных клеток, но обладает иными свойствами. Клетки расположены не параллельным пучком, как скелетные, а ветвятся, образуя единую сеть. Благодаря множеству клеточных контактов поступающий нервный импульс передается от одной клетки к другой, обеспечивая одновременное сокращение, а затем расслабление сердечной мышцы, что позволяет ей выполнять насосную функцию.
 … Клетки гладкой мышечной ткани не имеют поперечной исчерченности, они веретеновидные, одноядерные, их длина около 0, 1 мм. Этот вид ткани участвует в образовании стенок трубко-образных внутренних органов и сосудов (пищеварительного тракта, матки, мочевого пузыря, кровеносных и лимфатических сосудов).
… Клетки гладкой мышечной ткани не имеют поперечной исчерченности, они веретеновидные, одноядерные, их длина около 0, 1 мм. Этот вид ткани участвует в образовании стенок трубко-образных внутренних органов и сосудов (пищеварительного тракта, матки, мочевого пузыря, кровеносных и лимфатических сосудов).
 … Особенности гладкой мышечной ткани: — непроизвольность и небольшая сила сокращений, — способность к длительному тоническому сокращению, — меньшая утомляемость, — небольшая потребность в энергии и кислороде.
… Особенности гладкой мышечной ткани: — непроизвольность и небольшая сила сокращений, — способность к длительному тоническому сокращению, — меньшая утомляемость, — небольшая потребность в энергии и кислороде.


 … Нервная ткань, из которой построены головной и спинной мозг, нервные узлы и сплетения, периферические нервы, выполняет функции восприятия, переработки, хранения и передачи информации, поступающей как из окружающей среды, так и от органов самого организма. Деятельность нервной системы обеспечивает реакции организма на различные раздражители, регуляцию и координацию работы всех его органов.
… Нервная ткань, из которой построены головной и спинной мозг, нервные узлы и сплетения, периферические нервы, выполняет функции восприятия, переработки, хранения и передачи информации, поступающей как из окружающей среды, так и от органов самого организма. Деятельность нервной системы обеспечивает реакции организма на различные раздражители, регуляцию и координацию работы всех его органов.

 … Нейрон — состоит из тела и отростков двух видов. Тело нейрона представлено ядром и окружающей его областью цитоплазмы. Это метаболический центр нервной клетки; при его разрушении она погибает. Тела нейронов располагаются преимущественно в головном и спинном мозге, т. е. в центральной нервной системе (ЦНС), где их скопления образуют серое вещество мозга. Скопления тел нервных клеток за пределами ЦНС формируют нервные узлы, или ганглии.
… Нейрон — состоит из тела и отростков двух видов. Тело нейрона представлено ядром и окружающей его областью цитоплазмы. Это метаболический центр нервной клетки; при его разрушении она погибает. Тела нейронов располагаются преимущественно в головном и спинном мозге, т. е. в центральной нервной системе (ЦНС), где их скопления образуют серое вещество мозга. Скопления тел нервных клеток за пределами ЦНС формируют нервные узлы, или ганглии.
 Рисунок 2. Различные формы нейронов. а - нервная клетка с одним отростком; б - нервная клетка с двумя отростками; в - нервная клетка с большим количеством отростков. 1 - тело клетки; 2, 3 - отростки. Рисунок 3. Схема строения нейрона и нервного волокна 1 - тело нейрона; 2 - дендриты; 3 - аксон; 4 - коллатерали аксона; 5 - миелиновая оболочка нервного волокна; 6 - концевые разветвления нервного волокна. Стрелками показано направление распространения нервных импульсов (по Полякову).
Рисунок 2. Различные формы нейронов. а - нервная клетка с одним отростком; б - нервная клетка с двумя отростками; в - нервная клетка с большим количеством отростков. 1 - тело клетки; 2, 3 - отростки. Рисунок 3. Схема строения нейрона и нервного волокна 1 - тело нейрона; 2 - дендриты; 3 - аксон; 4 - коллатерали аксона; 5 - миелиновая оболочка нервного волокна; 6 - концевые разветвления нервного волокна. Стрелками показано направление распространения нервных импульсов (по Полякову).
 … Основными свойствами нервных клеток - являются возбудимость и проводимость. Возбудимость - это способность нервной ткани в ответ на раздражение приходить в состояние возбуждения.
… Основными свойствами нервных клеток - являются возбудимость и проводимость. Возбудимость - это способность нервной ткани в ответ на раздражение приходить в состояние возбуждения.
 … проводимость - способность передавать возбуждение в форме нервного импульса другой клетке (нервной, мышечной, железистой). Благодаря этим свойствам нервной ткани осуществляется восприятие, проведение и формирование ответной реакции организма на действие внешних и внутренних раздражителей.
… проводимость - способность передавать возбуждение в форме нервного импульса другой клетке (нервной, мышечной, железистой). Благодаря этим свойствам нервной ткани осуществляется восприятие, проведение и формирование ответной реакции организма на действие внешних и внутренних раздражителей.
ИСТОРИЯ. СРАВНИТЕЛЬНЫЕ ОСНОВЫ Г И СТОЛОГИИ
Гистология - учение о развитии, строении, жизнедеятельности и регенерации тканей животных организмов и организма человека.
Различают несколько уровней структурной организации организма:
1. молекулярный;
2. субклеточный:
3. клеточный;
4. тканевой;
5. органный;
6. системный;
7. организменный.
Разделами гистологии являются: цитология, эмбриология (учение о зародыше), общая гистология (наука о тканях), частная гистология (наука о гистофизиологии органов).
Гистология как наука имеет свои методы исследования:
1. Сравнительный или описательный;
2. Экспериментальный.
Эти методы основаны на применении различной оптической техники, поэтому можно выделить три этапа в развитии гистологии:
1. Домикроскопический. длился более 2000 лет (начало 400 лет до н.э.).
2. Микроскопический, длился около 300 лет. Начало связано с конструированием первых микроскопов и по усовершенствования современных. Первым микроскоп был создан в 1610 году (Г.Галилей). В 1665г. английский физик Р. Гук, рассматривая под микроскопом срез пробки, обнаружил, что она состоит из ячеек, напоминающих пчелиные соты. Эти образования Гук назвал клетками (лат. се11а - ячейка, клетка). Такое же строение Гук отметил в сердцевине бузины, камыша и некоторых других растений. Во второй половине XVII в. появились работы ряда микроскопистов: итальянца М. Мальпиги. англичанина Н. Грю, также обнаруживших ячеистое строение многих растительных объектов. Голландец А. Левенгук впервые обнаружил в воде одноклеточные организмы. Чешский ученый Я. Пуркине назвал полужидкое студенистое содержимое клетки протоплазмой. Английский ботаник Б. Броун обнаружил ядро. Немецкий зоолог Т. Шванн в 1839 г. обобщил все данные, которые были получены до него, и выдвинул основные положения клеточной теории. Р. Вирхов также внес большой вклад, развив и дополнив клеточную теорию; он написал труд "Целлюлярная патология".
Только в середине 19 века из микроскопической науки выделилась гистология. В этот же период гистология стала интенсивно развиваться в России. Сначала гистология преподавалась студентам на кафедре анатомии и физиологии. Поэтому первыми учеными-гистологами были анатомы, физиологи и эмбриологи. Первая кафедра гистологии была открыта в Московском университете в 1864 году профессором Овсянниковым. В это же время кафедра открылась в Военно-медицинской академии, возглавил ее Лавдовский. Только через 13 лет в Росси появился первый учебник Овсянникова и Лавдовского. Московскую кафедру гистологии возглавил А.И. Бабухин. Представители этих трех школ в своих исследованияхпроводили четкую гистофизиологическую позицию, т.е. не только описывали строение, но пытались объяснить закономерность строения, поэтому физиологическая направленность является приоритетной для отечественной гистологии.
Казанская школа морфологов известна своими трудами в области изучения нервной ткани, в том числе в.н.с. Арнштейн. Смирнов и Догель стали основателями этого направления. Поэтому в России многие вопросы о структуре органов и тканей стали рассматриваться с позиции нервной регуляции. Этому также способствовали работы Боткина. Павлова и Сеченова.
В начале 20 века в гистологии наиболее усиленно стали развиваться эволюционные подходы, основывавшиеся на работах Дарвина и Геккеля. Благодаря работам эмбриологов Вольфа, Нанлсра, Мечникова и Ковалевского, были продолжены искания в области эмбриологии и поддержаны эволюционные подходы.
Направленность советской гистологической школы была четкой в отношении клиники, поэтому большая часть гистологических работ была направлена на решение клинических задач.
3. Современный этап развития гистологии связан с более тонким изучением структур. Благодаря применению оптической, светооптической. Электронной микроскопии, гистохимических, количественных методов, цитофотометрии, были изучены органы на клеточном уровне, субклеточные структуры, молекулярные структуры. Задачи гистологии.
1. изучение структур на системном, органом, клеточном и молекулярном уровнях;
2. изучение физиологии структур всех уровней;
3. изучение закономерностей дифференцировки и регенерации:
4. изучение возрастных особенностей тканей и клеточных структур, включая закономерности эмбриогенеза:
5. изучение закономерностей адаптации структур всех уровней, в первую очередь связанных с проблемами экологии;
6. изучение закономерности нервной, эндокринной, иммунной регуляции.
ОСНОВЫ СРАВНИТЕЛЬНОЙ ЭМБРИОЛОГИИ
Эмбриология - это наука о развитии и строении зародыша человека.
Задачами медицинской эмбриологии являются изучение: прогенеза, этапов эмбриогенеза (от оплодотворения и до момента рождения), механизмов эмбриогенеза. Нарушений эмбрионального развития, возникновение причин нарушения развития, изучение критических периодов, разработка мер профилактики и изучение постнатального развития (т.е. развития после рождения) до периода полного становления всех органов и систем организма. В развитии выделяют: историческое развитие организма (филогенез) неиндивидуальное развитие организма (онтогенез). В онтогенезе выделяют эмбриогенез и постнатальное развитие. Наиболее ранним методом изучения эмбриологии является описательный метод, затем сравнительный и экспериментальный (это, прежде всего, искусственное оплодотворение) методы.
В эмбриогенезе выделяют периоды:
Оплодотворение:
Дробление: гистология эмбриология наука
Гаструляция:
Гистогенез;
Органогенез:
Системогенез:
Формирование организма в целом.
Оплодотворение является одним из этапов эмбриогенеза. В этом процессе участвует множество мужских половых клеток и одна женская. Но только ядро одного сперматозоида, сливаясь с яйцеклеткой, образует одноклеточный зародыш - зиготу, который несет материнский и отцовский наследственные генетические факторы. Оплодотворение в начале эволюции являлось внешним процессом. С выходом животных на сушу яйцеклетки стали выделяться во внешнюю воздушную среду. В связи с этим появлялись различные защитные оболочки: скорлуповая. белковая, желтковая. Через эти оболочки мужские половые клетки пройти не могут, поэтому оплодотворить такую яйцеклетку можно только до образования оболочек, т.е. внутри организма. Так возникает внутреннее оплодотворение.
Мужские половые клетки - сперматозоиды мало отличаются у различных видов животных и человека. Вырабатываются они в большом количестве, представляют собой мелкие и высокоподвижные клетки.
Женские половые клетки - яйцеклетки прошли более сложную эволюцию. Представляют собой крупные, малоподвижные клетки.
Вырабатываются в небольших количествах. Отличаются друг от друга количеством и распределением питательного желточного материала, а также размером.
Различают несколько типов яйцеклеток. Тип яйцеклетки зависит от длительности эмбрионального развития организма, от сложности его строения, от условий развития и от того, есть или нет личиночная стадия. Сначала появились первично изолеиитальные яйцеклетки. Они содержат мало желтка, и он равномерно распределен по всему объему клетки, диаметром около 100 мкм. Развитие животных с таким типом яйцеклетки идет в водной среде. Затем появляются телолецитальные яйцеклетки. У них возрастает содержание желтка, и он преимущественно локализуется на вегетативном полюсе. Также увеличивается размер яйцеклеток. Среди них выделяют умеренно телолецитальные (амфибии, рептилии) и резко телолецитальные (птицы) с очень высоким содержанием желтка. который полностью сосредоточен на вегетативном полюсе. Затем появляется вторично изолецитальная яйцеклетка (у высших млекопитающих и человека). Ее размер около 100 мкм. Содержит малое количество желтка, который равномерно распределен по всей цитоплазме. Вокруг клетки располагается блестящая оболочка, которая снаружи окружена эпителиальными клетками - "лучистый венец". Развитие таких организмов идет внутриутробно в материнском организме.
Причинами появления вторично изолецитальной яйцеклетки являются:
а} усложнение организма взрослой особи;
б) увеличение сроков эмбрионального развития;
в) изменение среды развития (матренискийорганизм);
г) исчезновение промежуточных личиночных стадий.
После оплодотворения начинается процесс дробления, в результате которого получается многоклеточный зародыш, имеющий у человека вид клеточного узелка - морулы. Затем в первичном узелке появляется, полость и образуется зародышевый пузырек или бластула. В процессе дробления зародыш в размерах не увеличивается, а возрастает только количество клеток (бластомеров), его составляющих.
Тип дробления определяется типом яйцеклетки. Различают:
У животных с первично изолецитальными яйцеклетками (ланцетник) дробление полное равномерное. При этом типе дробления бластомеры качественно не отличаются друг от друга, т.к. не происходит внутриклеточная дифференцировка;
У животных с умеренно телолецитальной яйцеклеткой (амфибии) дробление полное, но неравномерное;
У животных с резко телолецитальной яйцеклеткой (птицы) дробление неполное, дискоидальное:
У млекопитающих (и человека), имеющих вторичную изолецитальную яйцеклетку, дробление полное, неравномерное и асинхронное. При этом образуется нечетное количество бластомеров. Причем одни из них идут на образование зародыша, а другие - на образование провизорных органов (напр., трофобласта), которые создают условия для развития зародыша.
После образования бластулы начинается процесс гаструляции. На ранней стадии образуется двуслойный зародыш, а на поздней - трехслойный, который содержит наружный, средний и внутренний зародышевые листки (эктодерма, мезодерма и энтодерма) и комплекс осевых органов (хорда, нервная и кишечная трубка). Далее из зародышевых листков образуются ткани -гистогенез, а из тканей органы - органогенез.
Тип гаструляции ч определяется типом яйцеклетки. Выделяют: 1)впячивание, 2)эпиболию (обрастание), 3) иммиграцию, 4)деляминацию (расщепление). Для человека и высших млекопитающих характерен смешанный тип с преобладанием иммиграции и деляминации.
В процессе эмбриогенеза выделяют критические периоды, когда минимальные по силе факторы могут вызвать максимальное изменения в развитии. К таким периодам относятся:
Прогенез (образование половых клеток);
Процесс оплодотворения;
Дробление:
Гаструляция;
Имплантация (7-8 сутки);
Гистогенез;
Органогенез;
Системогенез:
Плацентарный (3-8 недель)
Процесс рождения ребенка.
Подобные документы
Гистология - наука о строении, развитии и жизнедеятельности тканей животных организмов и общих закономерностях тканевой организации; понятие цитологии и эмбриологии. Основные методы гистологического исследования; приготовление гистологического препарата.
презентация , добавлен 23.03.2013
Гистология как наука о происхождении, строении, функции и регенерации тканей живых организмов. Эволюционная эмбриология, развитие на примере млекопитающих. Критический период как период повышенной чувствительности организма к действию внешних факторов.
реферат , добавлен 18.01.2010
Изучение видов и функций различных тканей человека. Задачи науки гистологии, которая изучает строение тканей живых организмов. Особенности строения эпителиальной, нервной, мышечной ткани и тканей внутренней среды (соединительной, скелетной и жидкой).
презентация , добавлен 08.11.2013
История гистологии - раздела биологии, изучающего строение тканей живых организмов. Методы исследования в гистологии, приготовление гистологического препарата. Гистология ткани - филогенетически сложившейся системы клеток и неклеточных структур.
реферат , добавлен 07.01.2012
История зарождения гистологии как науки. Гистологические препараты и методы их исследования. Характеристика этапов приготовления гистологических препаратов: фиксация, проводка, заливка, резка, окрашивание и заключение срезов. Типология тканей человека.
презентация , добавлен 20.11.2014
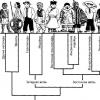
Факты о сходстве в строении человека и животных. Учение об эволюции, естественный отбор как его движущая сила. Доказательства происхождения человека от животных. Роль труда в процессе превращения обезьяны в человека. Факторы, вызвавшие антропоморфозы.
реферат , добавлен 02.10.2009
Общее описание эмбриологии человека. Образование внезародышевых оболочек. Описание стадий и этапов развития эмбриона. Характеристики поведения ребенка при токсических эффектах, последствий алкогольного синдрома. Синдром приобретенного иммунного дефицита.
реферат , добавлен 13.12.2008
Основные факты биографии Карла Эрнста фон Бэра - одного из основоположников эмбриологии и сравнительной анатомии, президента Русского энтомологического общества. Законы Бэра и его научные труды в области эмбриологии (закон зародышевого сходства).
презентация , добавлен 28.12.2015
Онтогенез как процесс формирования организмов с момента образования половых клеток и оплодотворения или отдельных групп клеток до завершения жизни. Исторические предпосылки и этапы развития эмбриологии как науки. Развитие одноклеточных организмов.
контрольная работа , добавлен 08.05.2011
Исследование отличительных свойств эпителиальных тканей. Изучение особенностей развития, строения и жизнедеятельности тканей организмов животных и человека. Анализ основных видов однослойного эпителия. Защитная и всасывающая функции эпителиальной ткани.
Лекция12 ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙ.
СИСТЕМА ЭПИТЕЛИАЛЬНЫХ ТКАНЕЙ
Планлекции
1. Введение, основные принципы организации тканей.
2. История изучения тканей и их классификация.
3. Общая характеристика эпителиальных тканей. Покровный эпителий. Строение и функции.
4. Железистый эпителий. Строение и функции.
Введение, основные принципы организации тканей. Ткань – фило-
генетически сложившаяся система клеток и неклеточных структур, объединённая, как правило, общностью происхождения, строения и специализированная на выполнении определенных функций (рис. 3.1 ).
Любую ткань необходимо рассматривать как частную систему по отношению к системе высшего ранга – организму. Ведущими элементами тка-
невой системы являются клетки, однако |
|||
большая роль в выполнении функций от- |
|||
водится межклеточному веществу. Кроме |
|||
клеток различают производные клеток – |
|||
симпласт, |
синцитий, |
постклеточные |
|
структуры. |
|||
Симпласт – многоядерная структура, |
|||
образованная |
при слиянии однотипных |
||
клеток, например, поперечно–полосатое |
|||
мышечное волокно. |
|||
Синцитий – структура, состоящая из |
|||
клеток, соединенных цитоплазматически- |
|||
ми мостиками. |
|||
К постклеточным структурам отно- |
|||
сятся эритроциты, тромбоциты, роговые |
|||
Рис. 3.1. Ткань – филогенетиче- |
чешуйки эпидермиса. |
||
ски сложившаяся система клеток и |
Межклеточное вещество (тканевый |
||
неклеточных структур: 1, 2 – клет- |
матрикс) подразделяют на основное веще- |
||
ки; 3 – межклеточное вещество |
ство и волокна. Основное вещество может |
||
(соединительно-тканные волокна) |
быть представлено гелем, золем или быть |
||
минерализовано. Среди волокон различа- |
|||
Цитология с основами гистологии. Конспект лекций |
|||
МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
ют три основных типа – ретикулярные, коллагеновые и эластические. Структуры тканевого матрикса построены из молекул, вырабатываемых и секретируемых клетками.
Клетки одинаковой морфофизиологической характеристики относятся к одному типу клеток, такое определение клеточного типа устраивало гистологов до определённого времени. В настоящее время наиболее правильным, строгим и современным определением понятия клеточного типа является следующее: клетки с идентичным набором разрешенных к экспрессии генов относятся к одному клеточному типу.
Клетки всегда находятся во взаимодействии друг с другом и с межклеточным веществом и формируют различные структурные объединения. Все межклеточные взаимодействия, как непосредственные, так и через межклеточное вещество, обеспечивают функционирование ткани как единой системы.
Как правило, ткань содержит совокупность клеточных форм, составляющих ту или иную линию дифференцировки – дифферон или гистогенетический ряд.
В диффероне последовательно различают: стволовые клетки → клеткипредшественницы → зрелые клетки, достигшие состо яния терминальной дифференцировки.
Стволовые клетки – самоподдерживающаяся популяция клеток, способных дифференцироваться в нескольких направлениях и формировать различные клеточные типы. Стволовые клетки обладают высокими пролиферативными потенциями, но, как правило, делятся редко.
Клетки-предшественницы по мере дифференцировки постепенно теряют свою пролиферативную потенцию. Наиболее ранняя стадия клетокпредшественниц – коммитированная, или полустволовая, клетка. При этом если клетка-предшественница может участвовать в образовании, например, трёх клеточных типов, то её непосредственный потомок может дифференцироваться только в двух направлениях и т.д.
Зрелая клетка – функционально активная клеточная форма, которой заканчивает гистогенетический ряд (дифферон).
Гистогенез (формирование ткани) – скоординированные в пространстве и времени процессы пролиферации, дифференцировки, детерминации, интеграции и функциональной адаптации.
Пролиферация протекает гиперплазией (увеличение числа клеток) и гипертрофией (увеличение массы клеток).
В ходе дифференцировки клетки накапливают те или иные внутриклеточные органоиды, образуя клеточные популяции. В нормальных условиях переход от более дифференцированного состояния к менее дифференцированному невозможен, то есть соблюдается принцип необратимости дифференцировки. Это свойство дифферона часто нарушается при новообразованиях (неоплазиях) – патологических разрастаниях клеток с нарушением кон-

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 12 ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙ. СИСТЕМА ЭПИТЕЛИАЛЬНЫХ ТКАНЕЙ
троля размножения и способности к построению тканевых и органных многоклеточных структур.
Детерминация – определение пути развития. Различают лабильную и стабильную степень детерминации, и чем выше дифференцировка, тем больше степень детерминации.
В гистогенезе все клетки интегрированы и создают вместе с межклеточным веществом морфофункциональные характеристики той или иной ткани.
Функциональная адаптация ткани – способность адекватно реагировать на изменения окружающей среды.
История изучения тканей и их классификация. Первая попытка сис-
тематизации тканей принадлежит французскому анатому Ксавье Биша, который в 1801г. выделял 21 разновидностъ тканей на макроскопическом уровне. В 1835–37 гг. Лейдиг и Келликер, основываясь на микроскопических исследованиях, предложили классификацию тканей, выделив 4 группы тканей: эпителиальные, соединительные, мышечные, нервные.
Изучая ткани позвоночных и беспозвоночных животных, А. А. Заварзин обратил внимание на сходное строение тканей, выполняющих одинаковую функцию, и создал теорию параллельных рядов тканевой эволюции. Суть теории заключается в том, что эволюция тканей шла параллельными рядами и в одном направлении – по пути увеличения числа клеточных форм и их специализации. Предложенная А. А. Заварзиным классификация тканей включает: систему пограничных тканей, систему тканей внутренней среды, систему мышечных тканей и ткани нервной системы.

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 12 ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙ. СИСТЕМА ЭПИТЕЛИАЛЬНЫХ ТКАНЕЙ
Хлопин Н. Г. создал теорию дивергентного развития тканей в фило- и онтогенезе. По Н. Г. Хлопину ткани в эволюции развивались путем расхождения признаков, и он дал генетическую классификацию тканей. Согласно Хлопину из восьми зачатков: энтодермы, целомической выстилки, энтомезенхимы, миотомов, хорды, кожной эктодермы, нейроэктодермы, прехордальной пластинки – образуются все виды тканей. Другими словами, в основу классификации тканей автор положил источники развития.
В настоящее время используется классификация тканей, основанная на морфологических особенностях тканей: система эпителиальных тканей, система соединительных тканей, система мышечных тканей, система нейральных тканей (рис.3.2 ).
Классификацияживотных тканей
Системанейтральных тканей
Собственно соединительная ткань
Рис. 3.2. Классификация животных тканей
Общая характеристика эпителиальных тканей. Покровный эпите-
лий. Строение и функции. Эпителиальная ткань занимает пограничное положение, отделяя организм от внешней среды, выстилает полости тела.
Характерными особенностями эпителиальных тканей являются:
оформление клеток в пласт (практически отсутствует межклеточное вещество);
гетерополярность клеток– наличие апикального и базального полюсов;
отсутствие собственных кровеносных сосудов;
наличие базальной мембраны, отделяющей эпителиальные клетки от рыхлой соединительной ткани, которая является источником питательных веществ для эпителия;
большая регенераторная способность;
![]()
МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 12 ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙ. СИСТЕМА ЭПИТЕЛИАЛЬНЫХ ТКАНЕЙ
эпителиоциты могут иметь органоиды специального назначения (реснички, жгутики, тонофибриллы).
Для системы эпителиальных тканей используется две классификации: гистогенетическая (по происхождению или источникам развития) и морфофункциональная (по строению и функции).
Согласно гистогенетической классификации выделяют:
1. Эпителии кожного типа (эктодермальные) – многослойный плоский ороговевающий и неороговевающий эпителий; эпителий слюнных, сальных, молочных и потовых желез; переходный эпителий мочеиспускательного канала; многорядный мерцательный эпителий воздухоносных путей; альвеолярный эпителий легких; эпителий щитовидной и паращитовидной железы, тимуса и аденогипофиза;
2. Эпителии кишечного типа (энтеродермальный) – однослойный призматический эпителий кишечного тракта; эпителий печени и поджелудочной железы;
3. Эпителий почечного типа (нефродермальный) – эпителий нефрона;
4. Эпителии целомического типа (целодермальный) – однослойный плоский эпителий серозных покровов (брюшины, плевры, околосердечной сумки); эпителий половых желез; эпителий коры надпочечников;
5. Эпителии нейроглиального типа – эпиндимный эпителий мозговых желудочков; эпителий мозговых оболочек; пигментный эпителий сетчатки глаза; обонятельный эпителий; глиальный эпителий органа слуха; вкусовой эпителий; эпителий передней камеры глаза; хромофобный эпителий мозгового слоя надпочечников; периневральный эпителий.
Для эпителиальных пластов принята морфофункциональная классификация, учитывающая количество слоев клеток (одно- и многослойные), рядность однослойного эпителия (однорядный, многорядный), форму клеток
и характер полярной дифференцировки.
Слойность определяется по наличию контакта с базальной мембраной. Если все клетки пласта находятся в контакте с базальной мембраной, такой эпителий определяется как однослойный. Если это условие не выполняется, эпителий – многослойный. Как правило, эктодермальные эпителии – многослойные, а энтодермальные – однослойные.
Рядность однослойных эпителиев отражает наличие или отсутствие в составе пласта клеток разной формы и типов. Рядность по сути определяется по расположению ядер клеток относительно базальной мембраны.
Форма клеток учитывает отношение их высоты к толщине. Различают плоский, кубический, цилиндрический (призматический) пласты эпителия. Морфофункциональная классификация однослойного эпителия:
1. Однослойный однорядный эпителий подразделяют: а) на однослойный плоский; б) однослойный кубический;
в) однослойный цилиндрический (призматический):
однослойный призматический каемчатый;

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 12 ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙ. СИСТЕМА ЭПИТЕЛИАЛЬНЫХ ТКАНЕЙ
однослойный призматический железистый;
однослойный призматический мерцательный.
2. Однослойный многорядный мерцательный эпителий.
3. Многослойный эпителий подразделяют:
а) на многослойный плоский неороговевающий; б) многослойный плоский ороговевающий; в) переходный.
В однослойном однорядном эпителии все клетки пласта без исключения непосредственно контактируют с базальной мембраной, имеют одинаковую высоту, поэтому ядра располагаются на одном уровне.
Однослойный плоский эпителий состоит из одного слоя резко уплощенных клеток полигональной формы, основание (ширина) клеток больше, чем высота (толщина). Такой тип эпителия выстилает серозные оболочки (брюшина, плевра, околосердечная сумка). В отношении эндотелия (клетки, выстилающие кровеносные и лимфатические сосуды, полости сердца) среди гистологов единого мнения нет: одни относят эндотелий к однослойному плоскому эпителию, другие – к соединительной ткани со специальными свойствами. Источники развития: эндотелий развивается из мезенхимы; однослойный плоский эпителий серозных покровов – из спланхнотомов (вентральная часть мезодермы). Такой эпителий имеет разграничительную функцию и уменьшает трение внутренних органов путем выделения серозной жидкости.
Однослойный кубический эпителий имеет вид куба, когда диаметр (ширина) клетки равен её высоте и встречается в выводных протоках экзокринных желез, в извитых почечных канальцах.
Однослойный призматический (цилиндрический) эпителий – это эпителий, у которого на срезе ширина клеток меньше, чем высота. В зависимости от особенностей строения и функции различают:
однослойный призматический железистый, имееется в желудке,
в канале шейки матки, специализирован на непрерывную выработку слизи;
однослойный призматический каемчатый, выстилает кишечник, на апикальной поверхности клеток имеется большое количество микроворсинок, специализированных на всасывание;
однослойный призматический реснитчатый, выстилает маточные трубы, на апикальной поверхности эпителиоциты имеют реснички.
Регенерация однослойного однорядного эпителия происходит за счет стволовых (камбиальных) клеток, равномерно разбросанных среди других дифференцированных клеток.
В однослойном многорядном мерцательном эпителии все клетки контактируют с базальной мембраной, но имеют разную высоту, и поэтому ядра располагаются на разных уровнях, т.е. в несколько рядов. Выстилает воздухоносные пути. В составе этого эпителия различают разновидности клеток:
короткие и длинные вставочные клетки;

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 12 ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙ. СИСТЕМА ЭПИТЕЛИАЛЬНЫХ ТКАНЕЙ
бокаловидные клетки, плохо воспринимают красители (в препарате – белые), вырабатывают слизь;
реснитчатые клетки, на апикальной поверхности имеют мерцательные реснички.
Функция однослойного многорядного мерцательного эпителия – очистка и увлажнение проходящего в легкие воздуха.
Многослойный эпителий состоит из нескольких слоев клеток, причем
с базальной мембраной контактирует только самый нижний ряд клеток. Многослойный плоский неороговевающий эпителий выстилает перед-
ний (ротовая полость, глотка, пищевод) и конечный отдел (анальный отдел прямой кишки) пищеварительной системы, роговицу. Состоит из следующих слоев:
1. Базальный слой – цилиндрической формы эпителиоциты со слабо базофильной цитоплазмой, часто с фигурой митоза; в небольшом количестве стволовые клетки для регенерации;
2. Шиповатый слой состоит из значительного количества слоев клеток шиповатой формы, клетки активно делятся;
3. Покровные клетки – плоские, стареющие клетки, не делятся, с п о- верхности постепенно слущиваются (рис. 3.3 ).
Рис. 3.3. Многослойный плоский неороговевающий эпителий роговицы глаза коровы: 1–клетки поверхностного слоя; 2–клетки среднего слоя; 3– клетки базального слоя; 4–базальная мембрана; 4–соединительная ткань

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 12 ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙ. СИСТЕМА ЭПИТЕЛИАЛЬНЫХ ТКАНЕЙ
Источник развития – эктодерма. Прехордальная пластинка – в составе энтодермы передней кишки. Функция многослойного плоского неороговеваюэпителия – механическая защита.
Многослойный плоский ороговевающий эпителий – это эпителий кожи. Развивается из эктодермы, зашищает от механических повреждений, лучевого, бактериального и химического воздействия, разграничивает организм и окружающую среду.
Состоит из слоев:
1. Базальный слой – во многом похож на аналогичный слой многослойного неороговевающего эпителия; дополнительно содержит до 10 % меланоцитов – отросчатых клеток с включениями меланина в цитоплазме, которые обеспечивают защиту от УФЛ; имеется небольшое количество клеток Меркеля (входят в состав механорецепторов); дендритические клетки с защитной функцией путем фагоцитоза; в эпителиоцитах содержатся тонофибриллы (органоид специального назначения), обеспечивающие прочность;
2. Шиповатый слой – эпителиоциты с шиповидными выростами; встречаются дендроциты и лимфоциты крови; эпителиоциты еще делятся;
3. Зернистый слой – несколько рядов вытянутых уплощенно-овальных клеток с базофильными гранулами кератогиалина (предшественник рогового вещества– кератина) в цитоплазме; клетки не делятся;
4. Блестящий слой – клетки полностью заполнены элаидином (образуется из кератина и продуктов распада тонофибрилл), отражающим и сильно преломляющим свет (под микроскопом границ клеток и ядер не видно);
5. Слой роговых чешуек – состоит из роговых пластинок из кератина, содержащих пузырьки с жиром и воздухом, кератосомы (соответствуют лизосомам). С поверхности чешуйки слущиваются (рис. 3.4 ).
Переходный эпителий выстилает полые органы, стенка которых способна сильно растягиваться (лоханка, мочеточники, мочевой пузырь).
Слои переходного эпителия:
1. Базальный слой – мелкие темные низкопризматические или кубические клетки – малодифференцированные и стволовые клетки, которые обеспечивают регенерацию;
2. Промежуточный слой – состоит из крупных грушевидных клеток
с узкой базальной частью, контактирует с базальной мембраной (стенка не растянута, поэтому эпителий утолщен); когда стенка органа растянута, грушевидные клетки уменьшаются по высоте и располагаются среди базальных клеток;
3. Покровные клетки – крупные куполообразные клетки; при растянутой стенке органа клетки уплощаются; клетки не делятся, постепенно слущиваются (рис. 3.5 ).

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 12 ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙ. СИСТЕМА ЭПИТЕЛИАЛЬНЫХ ТКАНЕЙ
Рис. 3.4. Многослойный плоский ороговевающий эпителий кожи пальца человека: 1 – роговой слой; 2 – блестящий слой; 3 – зернистый слой; 4 – шиповатый слой; 5 – базальный слой; 6 – базальная мембрана; 7 – выводной проток потовой железы; 8 – волокнистая соединительная ткань
Рис. 3.5. Переходный эпителий мочевого пузыря (нерастянутая стенка органа): 1 – клетки поверхностного плоского эпителия; 2 – клетки базального и промежуточного слоев эпителия; 3 – клетки на стадии амитоза и митоза; 4 – соединительная ткань

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 12 ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙ. СИСТЕМА ЭПИТЕЛИАЛЬНЫХ ТКАНЕЙ
Таким образом, строение переходного эпителия изменяется в зависимости от состояния органа: когда стенка не растянута, эпителий утолщен за счет «вытеснения» части клеток из базального слоя в промежуточный слой; при растянутой стенке толщина эпителия уменьшается за счет уплощения покровных клеток и перехода части клеток из промежуточного слоя в базальный. Источники развития: эпителий лоханки и мочеточника образуется из мезонефрального протока (производное сегментных ножек), эпителий мочевого пузыря – из энтодермы аллантоиса и энтодермы клоаки. Функция переходного эпителия – защитная.
Железистый эпителий. Строение и функции. Железистый эпителий специализирован на выработку секрета и образует железы. Классификация железистого эпителия по месту залегания:
Эндокринные железы не имеют выводящих протоков, и секрет выделяется непосредственно в кровь или лимфу. Эндокринные железы обильно снабжены кровеносными сосудами, которые доставляют в железу необходимые ингредиенты для синтеза гормонов и/или биологически активных веществ, оказывающих сильное регулирующее влияние на органы и системы даже в небольших дозах.
Экзокринные железы имеют выводящие протоки и выделяют секрет на поверхность эпителиев. Они образованы двумя видами эпителиоцитов, формирующих концевые (секреторные) отделы и выводящие пути (рис. 3.6 ).
Рис. 3.6. Строение и типы секреции экзокринных желез: А – эпителий; Б – волокнистая соединительная ткань; 1 – простая трубчатая железа; 2 – простая альвеолярная железа; 3 – сложная трубчатая железа; 4 – сложная альвеолярная железа; 5 – трубчатоальвеолярная железа; 6 – мерокриновый тип секреции; 7 – апокриновый тип секреции;
8 – голокриновый тип секреции (а – клетки росткового слоя; б – клетки распадающегося типа; в – секрет)

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 12 ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙ. СИСТЕМА ЭПИТЕЛИАЛЬНЫХ ТКАНЕЙ
Принципы классификации экзокринных желез: По строению выводящих протоков делятся:
на простые – выводящий проток не ветвится;
сложные – выводящий проток ветвится. По форме секреторных отделов делятся:
на альвеолярные – секреторный отдел в виде альвеолы, пузырька;
трубчатые – секреторный отдел в виде трубочки;
смешанные – трубчато-альвеолярные.
По соотношению выводящих протоков и секреторных отделов делятся:
на неразветвленные – в один выводящий проток открывается один секреторный отдел;
разветвленные – каждый секреторный отдел имеет свой выводящий проток, который сливается в один, общий выводящий, выделяющий секрет на поверхность эпителия.
По типу секреции делятся:
на мерокриновые –при выделении секрета целостность клеток не нарушается, что характерно для большинства желез;
апокриновые – секрет выделяется вместе с разрушением апикальной части клеток или верхушек микроворсинок (например, молочные железы);
голокриновые – секреция сопровождается полным разрушением железистых клеток (например, сальные железы кожи).
По локализации делятся:
на эндоэпителиальные – одноклеточная железа в толще покровного эпителия (например, бокаловидные клетки в эпителии кишечника и воздухоносных путях);
экзоэпителиальные железы – секреторный отдел лежит вне эпителия, в подлежащих тканях.
По характеру секрета: на белковые, слизистые, слизисто-белковые, липидные, белково-липидные и другие.
Секреция является сложным процессом, включающим четыре фазы:
1. Поглощение железистой клеткой исходных материалов для синтеза секрета (аминокислоты, липиды, углеводы, минеральные вещества и другие органические молекулы).
2. Синтез, созревание и накопление в железистых клетках секрета.
3. Выделение секрета.
4. Восстановление железистых клеток, имеющих апокриновый и голокриновый тип секреции.
В цитоплазме клеток железистого эпителия находятся органеллы, необходимые для синтеза секрета, имеющие разную степень развития в зависимости от характера секрета.
В железистом эпителии постоянно происходят процессы физиологической регенерации. В большинстве желез регенерация происходит путем деления специальных стволовых клеток, которые, дифференцируясь, превра-

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 12 ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙ. СИСТЕМА ЭПИТЕЛИАЛЬНЫХ ТКАНЕЙ
щаются в железистые клетки (клеточная регенерация). Отдельные железы (например, слюнные железы, поджелудочная железа) не имеют стволовых и малодифференцированных клеток, и в них происходит внутриклеточная регенерация изношенных органоидов при отсутствии способности к делению клеток.
Лекция13 Система соединительных тканей
Планлекции
1. Введение.
2. Собственно соединительная ткань. Типы. Клетки, межклеточное вещество, их строение и функции.
3. Соединительная ткань со специальными функциями.
4. Кровь как особый вид соединительной ткани.
Введение. Соединительные ткани – ткани мезенхимного генеза, имеющие широкое распространение, характеризующиеся разнообразием клеточных форм и хорошо развитым межклеточным веществом. Физико- химичес-кие особенности межклеточного вещества и строение его в значительной степени определяют функциональное значение разновидностей соединительных тканей (рис. 3.7 ).
Соединительные ткани выполняют механическую, опорную, формообразующую, защитную, пластическую и трофическую функции.
Рис. 3.7. Соединительные ткани

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Группу соединительных тканей составляют: собственно соединительные ткани, соединительные ткани со специальными свойствами и скелетные соединительные ткани.
Собственно соединительная ткань. Типы клеток, межклеточное вещество, их строение и функции. В основу классификации собственно соединительной ткани положен принцип соотношения клеток и степень упорядоченности расположения соединительно-тканных волокон межклеточного вещества. Собственно соединительную ткань подразделяют на волокнистые соединительные ткани (рыхлая и плотная), соединительные ткани со специальными свойствами. Плотная волокнистая соединительная ткань подразделяется на неоформленную и оформленную.
Рыхлая волокнистая соединительная ткань обнаруживается во всех органах, окружает и сопровождает кровеносные и лимфатические сосуды, располагается под базальной мембраной любого эпителия, образует прослойки и перегородки внутри всех паренхиматозных органов и слои в составе оболочек полых органов (рис. 3.8 ).
Рис. 3.8. Рыхлая волокнистая соединительная ткань: 1 – фибробласты; 2 – макрофаги; 3 – соединительно-тканное волокно
Клетки рыхлой волокнистой соединительной ткани представлены фибробластами, макрофагами, плазмоцитами, тканевыми базофилами (тучные клетки), адипоцитами, пигментоцитами (меланоциты), адвентициальными клетками, перицитами сосудов и лейкоцитами, которые мигрировали из крови. Клетки соединительной ткани гетерогенны по происхождению.

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 13 Система соединительных тканей
Фибробласты развиваются в эмбриогенезе из мезехимных клеток, после рождения – из стволовых клеток. Фибробластический дифферон представлен стволовыми клетками, фибробластами (полустволовые клетки-пред- шественники, малоспециализированные, дифференцированные – зрелые, активно функционирующие), фиброцитами (дефинитивные формы клеток), фиброкластами (клетки с большой фагоцитарной и гидролитической активностью), миофибробластами (сократительные клетки, имеющие общие черты с гладкомышечными клетками и содержащими актино-миозиновые комплексы). Основная функция активно функционирующих фибробластов состоит в синтезе компонентов межклеточного вещества в виде коллагена, эластина, фибронектина, гликозаминогликанов, протеогликанов, различных цитокинов.
Фибробласт – уплощенная клетка звездчатой формы, образует широкие клиновидные отростки; содержит крупное овальное ядро, несколько ядрышек, в цитоплазме располагается хорошо развитый шероховатый эндоплазматический ретикулум, аппарат Гольджи, много митохондрий, имеются лизосомы, секреторные пузырьки, гликоген, многочисленные микротрубочки и микрофиламенты (рис. 3.9 ).
Рис. 3.9. Фибробласт из кожи морской свинки
Макрофаги – вторая по численности группа клеток рыхлой неоформленной соединительной ткани. Макрофаги образуются из стволовой гемопоэтической клетки в результате дифференцировки моноцитов. Это профессиональные фагоциты, которые присутствуют во всех органах и тканях. Это очень мобильная популяция клеток, способная быстро перемещаться. Макрофаги подразделяют на резидентные (присутствуют в тканях в норме в отсутствии воспаления) и подвижные (популяция передвигающихся, так называемых «вызванных» макрофагов). Резидентные магрофаги подразделяют на свободные, имеющие округлую форму, и фиксированные – клетки звездча-
![]()
МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 13 Система соединительных тканей
той формы, прикрепляющиеся своими отростками к внеклеточному матриксу или другим клеткам.
Строение макрофагов зависит от их активности и локализации. Ядро макрофагов неправильной формы, в котором различаются глыбки хроматина. В цитоплазме присутствуют митохондрии, свободные рибосомы, хорошо выраженный комплекс Гольджи, гранулярная эндоплазматическая сеть, лизосомы, фаголизосомы, остаточные тельца. В лизосомах макрофагов идентифицируются миелопироксидаза, протеинкиназы, кислые гиролазы, лизоцим, катионные белки, лактоферрин, супероксиддисмутаза – антиоксидантный фермент, способствующий образованию Н2 О2 , ОН- , О2 2- . Плазмолемма макрофагов образует глубокие складки и длинные микровыросты, на поверхности плазмолеммы имеются рецепторы для опухолевых клеток, эритроцитов, Т - и В -лимфоцитов, антигенов и иммуноглобулинов. Под плазмолеммой
в большом количестве присутствуют актиновые микрофиламенты, микротрубочки, промежуточные филаменты, необходимые для миграции по механизму хемотаксиса и специфического и неспецифического фагоцитоза. Активированный макрофаг секретирует более 60 факторов.
Тучные клетки морфологически и функционально сходны с базофилами крови, хотя являются клетками различного клеточного типа. Тучные клетки происходят из предшественника в костном мозге, но окончательную дифференцировку претерпевают в соединительной ткани. Располагаются обычно вокруг кровеносных сосудов, много этих клеток в коже, в слизистой оболочке органов дыхательной и пищеварительной систем, матке, молочных железах, тимусе, миндалинах. Форма тканевых базофилов разнообразна – от округло-овальной до отростчатой с размерами от 4 до 14 мкм в ширину и до 20 мкм в длину. В цитоплазме очень много базофильных гранул, содержащих гепарин, гистамин и другие биологически активные вещества. Секретируемый клетками гепарин участвует в понижении свёртываемости крови, а гистамин участвует в воспалительных и аллергических реакциях. В целом тучные клетки регулируют местный гомеостаз.
Плазмоциты (плазматические клетки) образуются из иммунологически активированных В -лимфоцитов, синтезируют и секретируют Ig. По морфологии имеют сходство с лимфоцитами, хотя есть свои особенности. Ядро округлое или овальное, располагается несколько эксцентрично и содержит диспергированный гетерохроматин в виде пирамид, обращенных к центру острой вершиной и отграниченных друг от друга радиальными полосками. В цитоплазме хорошо развит аппарат Гольджи, гранулярная ЭПС. Цитоплазма базофильна, со светлым «двориком» около ядра. Величина плазмоцитов колеблется от 7 до 10 мкм.
Липоциты (адипоциты, жировые клетки) обладают способностью накапливать в больших количествах резервный жир, принимают участие в питании, энергообразовании и метаболизме воды. Адипоциты могут собираться
в группы и/или находиться по одиночке и, как правило, сопровождают кровеносные сосуды.

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 13 Система соединительных тканей
Зрелая жировая клетка обычно содержит одну большую каплю жира, которая оттесняет все внутриклеточные органоиды, в том числе и ядро, к периферии. В липоците обнаруживаются ферменты жирового обмена, которые обеспечивают высокий уровень метаболизма. Новые жировые клетки человека могут развиваться при усиленном питании из адвентициальных клеток.
Пигментоциты (меланоциты) лишь формально относятся к соединительной ткани, так как расположены в ней. В цитоплазме этих клеток содержится пигмент меланин. У людей черной и желтой рас пигментные клетки более распространены, чем у людей с белым цветом кожи.
Адвентициальные клетки – малодифференцированные клетки, располагаются рядом с кровеносными сосудами. Являются резервными клетками и могут дифференцироваться в другие клетки, например в фибробласты.
Перициты – отросчатые клетки, примыкающие снаружи к артериолам, венулам и капиллярам, участвуют в регуляции просвета капилляров, тем самым регулируют кровоснабжение окружающих тканей. Перициты имеют дисковидное ядро с небольшими углублениями и содержат обычный набор органелл. Функция перицитов связана с синтезом компонентов базальной мембраны капилляров. Кроме того, они контролируют пролиферацию эндотелиальных клеток при нормальном росте и регенерации. При заживлении ран и восстановлении сосудов перициты дифференцируются в гладкомышечные клетки.
Межклеточное вещество рыхлой волокнистой соединительной ткани состоит из основного вещества и волокон.
Различают коллагеновые, эластические и ретикулярные волокна. Коллагеновые волокна – главный компонент большинства соедини-
тельных тканей, содержит фибриллярный белок коллаген, представляющий спираль из трёх про-α-цепей. Под световым микроскопом имеют волнистый вид, длина спирали – 300 нм, диаметр – 1,5 нм. Молекулы коллагена синтезируются в фибробластах. Под поляризационным микроскопом коллагеновые волокна имеют продольную и поперечную исчерченность. Различают 13 типов коллагеновых волокон. В собственно соединительной ткани располагается коллаген первого типа. Сборка коллагеновых волокон происходит в межклеточном веществе, и имеется несколько уровней внеклеточной организации. Коллагеновые волокна не растягиваются, очень прочны на разрыв (6 кг/мм2 ), и это определяет их основную функцию, то есть обеспечивает механическую прочность ткани.
Эластические волокна – тонкие (d = 1–3 мкм), менее прочные (4–6 кг/см2 ), чем коллагеновые волокна,но зато очень эластичные волокна из белка эластина, синтезируются так же, как и коллагеновые волокна в фибробластах. Эти волокна исчерченностью не обладают, имеют прямой ход, часто разветвляются. Наличие эластических волокон в соединительной ткани определяет её эластичность и растяжимость. Форма волокон округлая или

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 13 Система соединительных тканей
уплощенная. В рыхлой волокнистой соединительной ткани эластические волокна анастомозируют друг с другом.
Ретикулярные волокна считаются разновидностью коллагеновых волокон и содержат повышенное количество углеводов, которые синтезируются ретикулярными клетками органов кроветворения. В отличие от коллагеновых волокон имеют меньший диаметр и сильно ветвятся, образуя трехмерную, петлистую сеть (ретикулум).
Клетки и волокна соединительной ткани заключены в аморфный компонент, или основное вещество, представляющее из себя гомогенную, гелеобразную, бесструктурную массу из макромолекул полисахаридов, связанных с тканевой жидкостью. Полисахариды представлены сульфатированными гликозаминогликанами, гепаринсульфатом, хондроэтинсульфатом, образующими комплексы с белками и гиалуроновой кислотой. Органическая часть основного вещества синтезируется в фибробластах, фиброцитах. Основное вещество, как коллоидная система, может переходить из состояния гель в состояние золь и наоборот и тем самым играет большую роль в рег у- ляции обмена веществ между кровью и другими тканями.
Плотная волокнистая соединительная ткань характеризуется большим количеством соединительно-тканных волокон коллагеновой природы, незначительным количеством аморфного вещества и клеточных компонентов. Со- единительно-тканные волокна могут располагаться упорядоченно и неупорядоченно, создавая оформленную и неоформленную плотную волокнистую соединительную ткань, соответственно. Клетки этой ткани представлены в подавляющем большинстве фибробластами и фиброцитами, в небольшом количестве встречаются макрофаги, тучные клетки, плазмоциты, малодифференцированные клетки. К оформленной плотной волокнистой соединительной ткани относятся сухожилия, связки,фасции, а к неоформленной – сетчатый слой дермы, капсулы паренхиматозных органов. В плотной волокнистой соединительной ткани между коллагеновыми волокнами встречаются прослойки рыхлой волокнистой соединительной ткани с кровеносными сосудами и нервными волокнами.
Соединительные ткани со специальными свойствами. К таким тка-
ням относят ретикулярную, жировую, слизистую и пигментную. Ретикулярная ткань составляет основу кроветворных органов, в не-
большом количестве имеется вокруг кровеносных сосудов. Состоит из ретикулярных клеток и межклеточного вещества, состоящего из основного вещества и ретикулярных волокон. Ретикулярные клетки, крупные с отростками
и оксифильной цитоплазмой, соединяясь друг с другом отростками и ретикулярными волокнами, образуют рыхлую сеть. Ретикулярные клетки способны к фагоцитозу, вырабатывают компоненты ретикулярных волокон. Ретикулярная ткань неплохо регенерирует за счет деления ретикулярных клеток
и выработки ими межклеточного вещества. Ретикулярная ткань выполняет опорно-механическую и трофическую функции для созревающих клеток крови, осуществляет фагоцитоз погибших клеток, инородных частиц, анти-

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 13 Система соединительных тканей
генов, создает специфическое микроокружение, определяет направление дифференцировки кроветворных клеток.
Жировая ткань – это скопления жировых клеток, располагающихся во многих органах. Условно различают две разновидности жировой ткани – бурую и белую, что связано с особенностями окраски клеток. Белая жировая ткань широко распространена в организме человека. Бурая жировая ткань встречается у новорожденных, а также у грызунов и зимоспящих животных.
Слизисто-студенистая ткань имеется только у эмбриона, располагается под кожей и в пупочном канатике. В этой ткани очень мало клеток, преобладает межклеточное вещество, богатое гиалуроновой кислотой, обеспечивающее высокий тургор данной ткани. Слизисто-студенистая ткань механически защищает нижележащие ткани и препятствует пережатию кровеносных сосудов пуповины.
Кровь как особый вид соединительной ткани. Кровь – жидкая со-
единительная ткань. Она состоит из жидкой части – плазмы и отдельных форменных элементов – эритроцитов, лейкоцитов и тромбоцитов. Форменные элементы крови образуются в кроветворных органах (в красном костном мозге, печени, селезёнке, лимфатических узлах).
В организме человека и животных кровь выполняет важные функции – дыхательную, трофическую, выделительную, защитную, гуморальную, участвует в терморегуляции.
Объем крови в теле человека с массой тела 70 кг составляет около 5–5,5 л. Кровь, межклеточное вещество и лимфа образуют внутреннюю среду организма, которая имеет постоянный состав. Это обеспечивает нормальный обмен веществ между клетками тканей и органов. Вместе с нервной и эндокринной системами кровь принимает участие в поддержании гомеостаза.
Плазма крови – это бесцветная жидкость, которая состоит на 90–93 % из воды и сухого вещества, в котором около 6,6–8,5 % принадлежит белкам и 1,5–3,5 % – органические и неорганические соединения.
Эритроциты, или красные кровяные тельца, у человека и млекопитающих представлены высокоспециализированными безъядерными клетками, содержащими гемоглобин для обеспечения транспортировки кислорода и углекислоты в организме. Кроме того, эритроциты участвуют в транспорте различных веществ и являются компонентом антиоксидантной системы организма.
Количество эритроцитов у женщин – 3,9–4,9 · 1012 /л, у мужчин – 4,0– 5,2 · 1012 /л, с диаметром 7–8 мкм. Эритроциты у человека и млекопитающих во взвешенном состоянии имеют форму двояковогнутого диска, такая конфигурация создаёт наибольшую площадь поверхности по отношению к объёму, что обеспечивает максимальный газообмен. Поверхность отдельного эритроцита приблизительно равна 125 мкм2 , а объем 90 мкм 3 , общая площадь п о- верхности циркулирующих в крови эритроцитов составляет около 3500–3700 м2 . В кров яное русло эритроциты выбрасываются из костного мозга в виде ретикулоцитов, имеющих в цитоплазме зернистость. Переход ретикулоцита

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 13 Система соединительных тканей
в эритроцит происходит в кровяном русле. Потенциальная продолжительность жизни эритроцитов составляет 100–120 дней. В сутки из кровотока удаляется 0,5–1,5 % общей массы эритроцитов и столько же вбрасывается.
Число эритроцитов у здоровых людей может варьироваться в зависимости от возраста, гормонального фона, психоэмоциональной и физической нагрузок, а также действия экологических факторов.
Лейкоциты (белые кровяные клетки) разнородны по морфологии и биологической роли. В одном литре крови взрослого человека содержится 3,8–9,8 · 109 лейкоцитов. Белые кровяные клетки имеют шаровидную форму, в цитоплазме которых находятся гранулы – специфические (вторичные) и азурофильные (лизосомы). В зависимости от типа гранул лейкоциты подразделяются на гранулоциты (зернистые) и агранулоциты (незернистые). Гранулоциты, к которым относятся нейтрофилы, эозирофилы, базофилы, содержат специфические и азурофильные гранулы и дольчатое сегментированное ядро разнообразной формы и называются полиморфноядерными лейкоцитами. Агранулоциты – моноциты и лимфоциты, содержат только азурофильные гранулы, имеют несегментированное ядро и называются мононуклеарными лейкоцитами. Содержание лейкоцитов в 1 мм3 крови и их соотношение (в %) представлено в табл. 3.1 .
Таблица 3.1 |
|||
Тип клетки |
Число клеток в 1 мм3 крови |
Соотношение, % |
|
Нейтрофилы |
|||
Эозинофилы |
|||
Базофилы |
|||
Моноциты |
|||
Лимфоциты |
|||
Полиморфноядерные лейкоциты образуются в костном мозге из кле- ток-предшественников, начало которым дают стволовые клетки. По мере созревания ядра в клетках появляются гранулы, типичные для каждого вида клеток. В кровотоке эти клетки перемещаются вдоль стенок капилляров в первую очередь за счет амебоидных движений. Нейтрофилы способны покидать внутреннее пространство сосуда и скапливаться в месте инфекции. Время жизни гранулоцитов около 10 дней, после чего они разрушаются в селезенке.
Нейтрофилы – наиболее многочисленные из лейкоцитов и составляют 40–75 % от общего количества лейкоцитов.
Диаметр нейтрофилов в мазке крови – 12–14 мкм. Большинство красителей окрашивает их ядро в фиолетовый цвет; ядро нейтрофилов периферической крови может иметь от одной до пяти долей. Цитоплазма окрашивается в розоватый цвет; под микроскопом в ней можно различить множество ин-

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 13 Система соединительных тканей
тенсивных розовых гранул. Количество митохондрий и органелл, необходимых для синтеза белка, минимально, и поэтому нейтрофилы не способны к продолжительному функционированию. У женщин примерно 1 % нейтрофилов несет половой хроматин (образованный одной из двух X-хромосом) – тельце в форме барабанной палочки, прикрепленное к одной из ядерных долей. Эти так называемые тельца Барра, позволяющие определять пол при исследовании образцов крови.
Главная функция нейтрофилов – фагоцитоз тканевых обломков и уничтожение микроорганизмов.
Базофилы составляют 0–1 % от общего числа лейкоцитов циркулирующей крови и размерами 10–12 мкм. Они имеют уплощенное ядро, которое состоит из чётко выраженных трёх долек, изогнутых в виде буквы S. В цитоплазме располагаются все виды органелл, свободные рибосомы, гликоген и цитоплазматические гранулы, окрашиваемые основными красителями в синий цвет. Активируемые базофилы могут покидать кровоток, выселяться в ткани и мигрировать к очагу воспаления, кроме того, участвовать в аллергических реакциях.
Моноциты – самые крупные лейкоциты с диаметром 15–20 мкм, количество их составляет 2–9 % от всех лейкоцитов циркулирующей крови. Они образуются в костном мозге. Крупное подковообразное, эксцентрично расположенное ядро моноцитов имеет пятнистый вид из-за неравномерно конденсированного хроматина. Цитоплазма при окраске голубовато-серая, содержит незначительное число включений, окрашивающихся красителем азуром в си- не-фиолетовый цвет. Моноциты образуются как в костном мозге, так и в селезенке и в лимфатических узлах. Их основная функция – фагоцитоз.
Лимфоциты – небольшие одноядерные клетки, составляют 20–45 % от общего числа лейкоцитов, циркулирующих в крови. Популяция лимфоцитов периферической крови неоднородна по размерам; их величина варьируется от 4,5 до 10 мкм. Принято выделять малые (4,5–6 мкм), средние (7–10 мкм) и большие лейкоциты (10–18 мкм) лейкоциты. Ядра клеток плотные и круглые, цитоплазма голубоватого цвета, с очень редкими гранулами. Несмотря на то, что лимфоциты выглядят морфологически однородно, они отчетливо

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 13 Система соединительных тканей
различаются по своим функциям и свойствам клеточной мембраны. Их делят на три большие категории: B -клетки, Т -клетки и NK -клетки.
B -лимфоциты составляют менее 10 % лимфоцитов крови, созревают у человека в костном мозге, после чего мигрируют в лимфоидные органы. Они служат предшественниками клеток, образующих антитела (плазматические клетки). Для того, чтобы B -клетки трансформировались в плазматические, необходимо присутствие Т -клеток.
Созревание Т -клеток начинается в костном мозге, где образуются протимоциты, которые затем мигрируют в тимус (в илочковую железу) – орган, расположенный в грудной клетке за грудиной. Там они дифференцируются в Т -лимфоциты – весьма неоднородную популяцию клеток иммунной системы, выполняющих различные функции. Т -лимфоциты составляют большинство лимфоцитов крови, на их долю приходится 80 % и более. Главная фун к- ция – участие в клеточном и гуморальном иммунитете. Они синтезируют факторы активации макрофагов, факторы роста B -клеток и интерфероны. Есть среди Т -клеток индукторные (хелперные) клетки, которые стимулируют образование B -клетками антител. Есть и клетки-супрессоры, которые подавляют функции B -клеток и синтезируют фактор роста Т -клеток – интерлейкин- 2 (один из лимфокинов).
NK -клетки отличаются от B - и Т -клеток тем, что у них нет поверхностных детерминант. Некоторые из них служат «естественными киллерами», т. е. убивают раковые клетки и клетки, зараженные вирусом. Однако в целом роль NK -клеток неясна.
Тромбоциты представляют собой бесцветные безъядерные тельца сферической, овальной или палочкообразной формы диаметром 2–4 мкм. В норме содержание тромбоцитов в периферической крови составляет 200000– 400000 на 1 мм3 . Продолжительность их жизни – 8–10 дней. Стандартными красителями (азур-эозин) они окрашиваются в однородный бледно-розовый цвет. С помощью электронной микроскопии показано, что тромбоциты являются фрагментами цитоплазмы очень крупных клеток мегакариоцитов, присутствующих в костном мозге. Мегакариоциты происходят из потомков тех же стволовых клеток, которые дают начало эритроцитам и лейкоцитам. Тромбоциты играют ключевую роль в свертывании крови. Кроме того, в последнее время отмечено участие тромбоцитов в аллергических реакциях и восстановлении целостности сосудов.

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция14 Скелетные ткани. Система мышечных тканей.
Гладкая мышечная ткань
Планлекции
1. Общая характеристика хрящевой ткани. Клетки хрящевой ткани.
2. Межклеточное вещество хрящевой ткани.
3. Общая характеристика костной ткани. Клетки костной ткани. Костный матрикс.
4. Типы костной ткани и их строение. Остеогенез.
5. Общая характеристика мышечной ткани. Гладкая мышечная ткань. Ее строение.
Общая характеристика хрящевой ткани. Клетки хрящевой ткани.
К скелетным типам тканей внутренней среды относятся хрящевая и костная ткани, выполняющие опорную, защитную, механическую функции и принимающие участие в водно-солевом обмене. Хрящевая ткань состоит клеток – хондроцитов и хондробластов, а также из большого количества межклеточного вещества, отличающегося прочностью и упругостью. Плотное межклеточное вещество (матрикс) представлено аморфным и волокнистым компонентами. Хрящевая ткань содержит 10–12 % органических соединений, 4–6 % минеральных солей, и остальную долю составляет вода. Хрящевая ткань, как
и покровный эпителий, не содержит кровеносных сосудов. При развитии хрящей ткани из мезенхимы мезенхима уплотняется, при этом клетки теряют свои отростки, усиленно пролиферируют и образуют скелетогенный зачаток, из которого хонрогенные клетки дифференцируются в хондробласты. Молодые хондробласты начинают создавать тонкие прослойки межклеточного вещества и образуют первичную хрящевую ткань или прехондриальную ткань. По периферии хрящевой закладки на границе с мезенхимой возникает надхрящница или перихондрий, состоящий из наружного соединительного и внутреннего хондрогенного слоев. В хондрогенной зоне клетки интенсивно делятся и наслаиваются на уже имеющийся хрящ по его периферии. Так происходит аппозиционный рост хряща. Хрящевые клетки, лежащие в толще молодого хряща, ещё какое-то время делятся, что увеличивает массу хряща изнутри. Такой рост хряща называется интерстициальным.
Хондробласты располагаются одиночно по периферии хрящевой ткани. Представляют собой вытянутые уплощенные клетки с базофильной цитоплазмой, содержащей хорошо развитую зернистую эндоплазматическую сеть
и аппарат Гольджи. Эти клетки синтезируют компоненты межклеточного вещества, выделяют их в матрикс и постепенно дифференцируются в дефинитивные клетки хрящевой ткани – хондроциты. Хондробласты обладают способностью митотического деления (рис. 3.10 ).

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Рис. 3.10. Хондробласты различной степени дифференцировки
Хондроциты по степени зрелости, морфологии и функции подразделяются на клетки I, II и III типа.
Рис. 3.11. Изогенные группы хондроцитов
Все разновидности хондроцитов локализуются в более глубоких слоях хрящевой ткани в особых полостях – лакунах, являющихся продуктом жизнедеятельности хрящевых клеток. Молодые хондроциты (I типа) митотически делятся, однако дочерние клетки оказываются в одной лакуне и образуют группу клеток – изогенную группу (клон). Изогенная группа является общей структурно-функциональной единицей хрящевой ткани. Расположение хондроцитов в изогенных группах в разных хрящевых тканях неодина-
ково (рис. 3.11 ).
Межклеточное вещество хрящевой ткани. Содержит до 75 % воды,
что позволяет веществам с низкой молекулярной массой из сосудов надхрящницы диффундировать в матрикс и осуществлять питание хондроцитов. Важное значение для обеспечения прочности и упругости хряща имеют белки хрящевого матрикса. Функционально наиболее значимы коллагены, протеогликаны и хондронектин. Коллаген II типа, образующий коллагеновые

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 14 Скелетные ткани. Система мышечных тканей. Гладкая мышечная ткань
волокна, составляет до 40 % сухого веса хряща. Коллаген IX сшивает коллагеновые волокна, а такжеα2 - цепь ковалентно связывает хондроитинсульфат. Содержание коллагена IX в хряще в пять раз меньше, чем коллагена II. Аморфное вещество содержит, главным образом, сульфатированные гликозоаминогликаны, и прежде всего хондроитинсуфаты, а также протеогликаны. Гликозоаминогликаны связывают большое количество воды и обуславливают плотность межклеточного вещества. Кроме того, в аморфном веществе содержится значительное количество минеральных веществ, не образующих кристаллы. Хондронектин, контролируя консистенцию матрикса, важен для развития хряща и поддержания его структуры.
В зависимости от строения межклеточного вещества хрящевые ткани подразделяют на гиалиновую, эластическую и волокнистую.
Общая характеристика костной ткани. Клетки костной ткани. Ко-
стный матрикс. Костная ткань – особая форма соединительной ткани, в которой около 70 % от её сухого вещества приходится на неорганические соединения (рис. 3.12 ). У позвоночных в постнатальном периоде развития организма из такой ткани сформированы кости скелета. Остеогенные клетки происходят из мезенхимы, имеют веретеновидную форму. При высоком рО2 остеогенные клетки дифференцируются в остеобласты, а при низком рО2 – в хондрогенные клетки.
Рис. 3.12. Костная ткань
Остеобласты представляют из себя популяцию неделящихся отростчатых клеток, имеющих кубическую, полиганальную или цилиндрическую форму. Ядро расположено эксцентрично, цитоплазма резко базофильна. Размеры тела остеобластов составляют 15–20 мкм. Основная функция остеобластов связана с синтезом и секрецией костного матрикса. Остеобласты синтезируют белок коллаген и гликозоаминогликаны. За счет этих компонентов формируется органический матрикс костной ткани. Затем эти же клетки
![]()
МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 14 Скелетные ткани. Система мышечных тканей. Гладкая мышечная ткань
обеспечивают минерализацию межклеточного вещества посредством выделения солей кальция. Постепенно, выделяя межклеточное вещество, они как бы замуровываются и превращаются в остеоциты. При этом внутриклеточные органеллы в значительной степени редуцируются, синтетическая и секреторная активность снижается и сохраняется функциональная активность, свойственная остеоцитам. Остеобласты, локализующиеся в камбиальном слое надкостницы, находятся в неактивном состоянии, синтетические и транспортные органеллы слабо развиты. При травмах и переломах костей в цитоплазме остеобластов быстро развивается зернистая эндоплазматическая сеть, аппарат Гольджи и происходит активный синтез и выделение коллагена и гликозоаминогликанов, для формирования органического матрикса создается костная мозоль, а затем идет формирование дефинитивной костной ткани. Таким способом за счет деятельности остеобластов надкостницы происходит регенерация костей при их повреждении.
Остеоциты – зрелые неделящиеся клетки, расположенные в костных полостях или лагунах, повторяющих контуры остеоцита. Длина полостей варьирует от 22 до 55 мкм, а ширина – от 6 до 14 мкм. Эти клетки имеют о т- ростки, которые находятся в костных канальцах. В цитоплазме локализовано крупное компактное ядро, органеллы развиты слабо. Многочисленные костные канальцы, анастомозируя между собой, пронизывают всю костную ткань, сообщаясь с периваскулярными пространствами, и образуют дренажную систему костной ткани. В этой дренажной системе содержится тканевая жидкость, посредством которой обеспечивается обмен веществ не только между клетками и тканевой жидкостью, но и с межклеточным веществом. В ядре преобладает гетерохроматин. Остеоциты обладают незначительной функциональной активностью, которая заключается в поддержании обмена веществ между клетками и межклеточным веществом (рис. 3.13 ).
Остеокласты – костеразрушающие
клетки, в сформированной костной ткани |
|||
они отсутствуют, но содержатся в надко- |
|||
стнице и в местах разрушения и пер |
|||
стройки костной ткани. Остеокласты – |
|||
крупные (диаметр 90 мкм и более) мно- |
|||
гоядерные клетки, дифференцируются |
|||
из моноцитов и относятся к системе мо- |
|||
нонуклеарных фагоцитов. Поскольку в |
|||
онтогенезе непрерывно осуществляются |
|||
локальные процессы перестройки кост- |
|||
ной ткани, то в этих местах обязательно |
|||
Рис. 3.13. Костные клетки |
присутствуют и остеокласты. В процессе |
||
эмбрионального остеогистогенеза |
|||
клетки играют важную роль и определяются в большом количестве. Остеокласты имеют характерную морфологию:
эти клетки являются многоядерными (3 и более ядер);

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 14 Скелетные ткани. Система мышечных тканей. Гладкая мышечная ткань
это довольно крупные клетки (диаметром около 90 мкм);
они имеют характерную овальную форму, но часть ее, прилежащая
к костной ткани, является плоской.
При этом в плоской части выделяют две зоны. Первая зона, наиболее обширная, богатая цитоплазматическими выростами, так называемая гофрированная каемка, является областью абсорбции и секреции гидролитических ферментов. Вторая зона – зона плотного контакта остеокласта к костной поверхности, окружая первую, она как бы герметизирует область действия ферментов. В этой зоне цитоплазма светлая, содержит мало органелл, большое количество микрофиламентов актиновой природы.
В цитоплазме клетки, под ядрами, располагаются многочисленные лизосомы и вакуоли разной величины. Функциональная активность остеокласта проявляется следующим образом: в гофрированной зоне из остекласта выделяется большое количество Н+ и СI- , что создает и поддерживает в замкнутом пространстве лакуны кислую среду, а фермент карбоангидраза II способствует образованию угольной кислоты. Выделяющаяся угольная кислота вызывает деминерализацию костной ткани, а протеолитические ферменты разрушают органический матрикс межклеточного вещества. Фрагменты коллагеновых волокон фагоцитируются остеокластами и разрушаются внутриклеточно. Посредством этих механизмов происходит резорбция (разрушение) костной ткани, и потому остеокласты обычно локализуются в углублениях костной ткани. После разрушения костной ткани за счет деятельности остеобластов, выселяющихся из соединительной ткани сосудов, происходит построение новой костной ткани.
Костный матрикс составляет 50 % сухого веса кости и состоит из неорганической (50 %) и органической (25 %) частей и воды (25 %). Неорганическая часть в значительном количестве содержит гидроксиапатит, который вместе с другими неорганическими веществами образуют кристаллы. Остеонектин связывает коллаген с кристаллами гидроксиапатита. В состав неорганической составляющей костного матрикса также входят бикарбонаты, цитраты, фториды, соли Мg2+ , К+ , Nа+ . Органическая часть костного матрикса представлена коллагеном типа I – 90–95 % и коллагеном типа V, неколлагеновыми белками – остеонектином, остеокальцином, сиалопротеинами, протеолипидами, а также глюкозамингликанами. Остеонектин поддерживает
в присутствии коллагена осаждение Са 2+ и РО4 3- . Остеокальцин участвует
в процессе кальцификации и служит маркёром для оценки активности кости.
Типы костной ткани и их строение. Остеогенез. Существует два ос-
новных типа костной ткани – грубоволокнистая (ретикулофиброзная) и пластинчатая. Эти разновидности костной ткани различаются структурными, физическими свойствами, которые коренятся, главным образом, в строении межклеточного вещества.
Грубоволокнистая костная ткань встречается главным образом у зародышей. У взрослых её можно встретить на месте заросших черепных швов и в местах прикреплений сухожилий к костям. Коллагеновые волокна образу-

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 14 Скелетные ткани. Система мышечных тканей. Гладкая мышечная ткань
ют беспорядочно расположенные толстые пучки. В основном веществе ретикулофиброзной костной ткани находятся удлиненно-овальные лакуны, с длинными анастомозирующими канальцами, повторяющими форму остеоцитов.
Пластинчатая костная ткань относится к зрелой (вторичной) костной ткани. Она образована костными пластинками и является наиболее распространенной разновидностью костной ткани во взрослом организме. Пластинчатая костная ткань формирует губчатое и компактное вещество кости. Губчатое вещество представляет из себя переплетающиеся траберукулы, полости между которыми заполнены костным мозгом. Трабекула состоит из костных пластинок и снаружи окружена одним слоем остеобластов. Трабекулы расположены соответственно направлению сил сжатия и растяжения. Губчатое вещество заполняет эпифизы длинных трубчатых костей и образует внутреннее содержимое коротких и плоских костей скелета. Основная масса компактного вещества состоит из остеонов, образует диафизы длинных трубчатых костей и слоем различной толщины покрывает все остальные кости скелета.
Костная пластина – слой костного матрикса толщиной 3–7 мкм, с о- стоящий из костных пластинок, образованных костными клетками и минеральным аморфным веществом с коллагеновыми волокнами. В толще пластинки в костных канальцах проходят отростки остеоцитов. Коллагеновые волокна в пределах пластинки ориентированы упорядоченно и лежат под углом к волокнам соседней пластинки, благодаря чему достигается большая прочность пластинчатой костной ткани.
Остеон, или хаверсова система, построена из 4–20 костных пластинок.
В центре остеона расположен хаверсовый канал (канал остеона), заполненный рыхлой волокнистой соединительной тканью с кровеносными сосудами
и нервными волокнами. Каналы Фолькмана связывают каналы остеона между собой, а также с сосудами и нервами надкостницы. Снаружи остеон образует спайную линию (линия цементации), отделяющую его от фрагментов старых остеонов. В ходе образования остеона находящиеся в непосредственной близости от сосуда хаверсового канала остеогенные клетки дифференцируются в остеобласты. Снаружи располагается сформированный остеобластами слой остеоида (неминерализованный органический костный матрикс).
В дальнейшем остеоид минерализуется и остеобласты дифференцируются
в остеоциты. Следующий концентрический слой возникает изнутри подобным образом. По наружной поверхности остеоида на границе с минерализованным костным матриксом проходит фронт обызвествления, где начинается процесс отложения минеральных солей.
Различают два способа остеогистогенеза:
1. Развитие непосредственно из мезенхимы – прямой (внутримембранный) остеогистогенез;
2. Развитие из мезенхимы через стадию хряща – непрямой (энхондриальный) остеогистогенез.

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 14 Скелетные ткани. Система мышечных тканей. Гладкая мышечная ткань
Посредством прямого остеогистогенеза развивается небольшое количество костей (плоские кости). В участках мезенхимы, содержащих кровеносные сосуды, группы мезенхимных клеток формируют первичные центры окостенения. Мезенхимные клетки дифференцируются в остеобласты, которые начинают вырабатывать остеоид. Остеоид минерализуется, остеобласты переходят в остеоциты, замурованные в лакунах минерализованного костного матрикса. Сформировавшаяся незрелая ткань существует в форме трабекул. Отдельные трабекулы различных участков растут и объединяются друг с другом. Анастомозирующая сеть костных трабекул формирует губчатое вещество, которое замещается пластинчатой костной тканью.
Непрямой остеогистогенез начинается со 2-го месяца эмбриогенеза. Вначале в мезенхиме за счет деятельности хондробластов закладывается хрящевая модель будущей кости из гиалиновой хрящевой ткани. Морфогенетические белки кости индуцируют энхондральный остеогенез. Затем происходит замена хрящевой ткани костной, вначале в диафизах, а затем в эпифизах.
Окостенение в диафизе осуществляется двумя способам: перихондрально и энхондрально.
Вначале в области диафиза хрящевой закладки кости из надхрящницы выселяются остеобласты и образуют ретикулофиброзную костную ткань, которая в виде манжетки охватывает по периферии хрящевую ткань. В результате этого надхрящница превращается в надкостницу. Такой способ образования костной ткани называется перихондральным. После образования костной манжетки нарушается трофика глубоких частей гиалинового хряща в области диафиза, в результате чего здесь происходит отложение солей кальция – омеление хряща. Затем под индуктивным влиянием обызвествленного хряща, в эту зону из надкостницы через отверстие в костной манжетке прорастают кровеносные сосуды, в адвентиции которых содержатся остеокласты и остеобласты.
Остеокласты разрушают омелевший хрящ, за счет деятельности остеобластов формируется пластинчатая костная ткань в виде первичных остеонов, которые характеризуются широким просветом (каналом) в центре и нечеткими границами между пластинками. Такой способ образования костной ткани в глубине хрящевой ткани и носит название энхондрального.
Одновременно с энхондральным окостенением происходит перестройка грубоволокнистой костной манжетки в пластинчатую костную ткань, составляющую наружный слой генеральных пластин. В результате перихондрального и энхондрального окостенения хрящевая ткань в области диафиза замещается костной. При этом формируется полость диафиза, заполняющаяся вначале красным костным мозгом, сменяющимся затем на желтый костный мозг.
Эпифизы трубчатых костей и губчатые кости развиваются только энхондрально. Вначале в глубоких частях хрящевой ткани эпифиза отмечается омеление. Затем туда проникают сосуды с остекластами и остеобластами и за

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 14 Скелетные ткани. Система мышечных тканей. Гладкая мышечная ткань
счет их деятельности происходит замена хрящевой ткани пластинчатой в виде трабекул. Периферическая часть хрящевой ткани сохраняется в виде суставного хряща. Между диафизом и эпифизом длительное время сохраняется хрящевая ткань – метаэпифизарная пластинка, за счет постоянного размножения клеток метафизарной пластинки происходит рост костей в длину.
В метаэпифизарной пластинке выделяют три зоны клеток:
1. Пограничная зона;
2. Зона столбчатых клеток;
3. Зона пузырчатых клеток.
Примерно к двадцати годам метаэпифизарные пластинки редуцируются, происходит синостозирование эпифизов и диафиза, после чего рост костей в длину прекращается. В процессе развития костей за счет деятельности остеобластов надкостницы происходит рост костей в толщину.
Регенерация костей после их повреждения и переломов осуществляется за счет деятельности остеобластов надкостницы. Перестройка костной ткани осуществляется постоянно на протяжении всего онтогенеза: одни остеоны или их части разрушаются, другие – образуются. Таким образом, костная ткань является динамической системой.
Общая характеристика мышечной ткани. Гладкая мышечная ткань. Ее строение. Мышечная ткань осуществляет двигательные функции организма. Во всех сократительных элементах мышечной ткани функционирует актиномиозиновый хемомеханический преобразователь. Кроме актина
и миозина в процессе сокращения – расслабления мышечных элементов участвуют регуляторные белки и Ca 2+ . У части гистологических элементов мышечной ткани видны сократительные единицы – саркомеры, которые выявляют поперечно-полосатую исчерченность ткани, а у другой части мышечной ткани исчерченность отсутствует. Это обстоятельство позволяет различать два типа мышечной ткани: поперечно-полосатую (исчерченную) мышечную ткань, которая в свою очередь подразделяется на скелетную и сердечную,
и гладкую (неисчерченную) мышечную ткань (рис. 3.14 ).
Сокращение гладких мышц инициируется нервными импульсами, некоторыми гормонами и не зависит от воли человека, так как их тонус не контролируется нашим сознанием. К гладким мышцам относятся мышцы внутренних органов, системы пищеварения, стенок кровеносных сосудов, кожи и матки, обеспечивая их сокращение и расслабление.

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 14 Скелетные ткани. Система мышечных тканей. Гладкая мышечная ткань
Рис. 3.14. Виды мышечной ткани: 1 – поперечно-полосатая скелетная; 2 – попереч- но-полосатая сердечная; 3 – гладкая
Функционально сердечная мышца отличается от скелетной и занимает промежуточное положение между гладкими и скелетными мышцами. Сердечная мышца сокращается ритмично с последовательно меняющимися циклами сокращения (систола) и расслабления (диастола) независимо от воли человека, непроизвольно. Ее сокращение регулируется гормонами.
Скелетная мускулатура относится к поперечно-полосатой мускулатуре и обеспечивает перемещение человека в пространстве. В организме взрослого мужчины скелетная мускулатура составляет более 40 % общей массы, у пожилых людей – около 30 %, у детей – около 25 %, у женщин – меньше. Проявление различных двигательных качеств человека, особенно силы и скорости, зависит от морфологического строения мышц, особенностей протекания биохимических процессов в них, а также от регуляторного воздействия нервной системы.
Гладкая мышечная ткань развивается из мезенхимы. Она составляет двигательный аппарат внутренних органов, кровеносных и лимфатических сосудов. Ее сокращения имеют медленный, тонический характер. Структурной единицей гладкой мышечной ткани является клетка удлиненной веретенообразной формы - гладкий миоцит. Она покрыта плазмолеммой, к которой снаружи примыкает базальная мембрана и соединительно-тканные волокна. Внутри клетки в ее центре, в миоплазме, имеется вытянутой формы ядро, вокруг которого расположены митохондрии и другие органеллы.
В миоплазме миоцитов под электронным микроскопом обнаружены сократительные белковые нити - миофиламенты. Различают миофиламенты актиновые, миозиновые и промежуточные. Актиновые и миозиновые миофиламенты обеспечивают сам акт сокращения, а промежуточные предохраняют гладкие миоциты от их избыточного расширения при укорочении. Миофиламенты гладких миоцитов не образуют дисков, поэтому эти клетки не имеют

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 14 Скелетные ткани. Система мышечных тканей. Гладкая мышечная ткань
поперечной исчерченности и получили название гладких, неисчерченных. Гладкие миоциты хорошо регенерируют. Они делятся митозом, могут развиваться из малодифференцированных соединительно-тканных клеток, способны к гипертрофии. Между клетками располагается опорная строма гладкой мышечной ткани - коллагеновые и эластические волокна, образующие плотные сети вокруг каждой клетки.
Гладкие мышечные клетки синтезируют сами волокна этой стромы. Особенности гладкой мышечной ткани: непроизвольность и небольшая
сила сокращений, способность к длительному тоническому сокращению, меньшая утомляемость, небольшая потребность в энергии и кислороде.
Лекция15 Поперечно-полосатая мышечная ткань.
Нервная ткань
Планлекции
1. Поперечно-полосатая мышечная ткань. Особенности строения и функции.
2. Скелетная поперечно-полосатая мышечная ткань. Строение саркомера и структура поперечно-полосатого мышечного волокна.
3. Актин и миозин – основные сократительные белки.
4. Сердечная поперечно-полосатая мышечная ткань.
5. Нервная ткань. Общая характеристика.
6. Нейроциты.
7. Нейроглия.
Поперечно–полосатая мышечная ткань. Особенности строения и функции. Поперечно–полосатая мышечная ткань подразделяется на скелетную и сердечную. Источником развития элементов скелетной поперечно– полосатой мышечной ткани являются клетки миотомов. Одни из них дифференцируются на месте, другие же мигрируют из миотомов в мезенхиму. В мезенхиме эти клетки детерменированы в направлении развития элементов мышечной ткани. Морфологически они ничем не отличаются от клеток мезенхимы. Их дифференцировка продолжается в местах закладки будущих мышц.
При этом возникают две линии дифференцировки. Клетки одной из них сливаются, образуя симпластические структуры – мышечные трубочки (миотубы). В них происходит дифференцировка специальных органелл – миофибрилл, которые сначала располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра из центральной области смещаются к периферии – формируется миосимпласт. Клетки другой линии остаются

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
самостоятельными, дифференцируются в миосателлитоциты, которым отводится роль восстановления мышечной ткани при повреждении.
Скелетная поперечно–полосатая мышечная ткань. Строение саркомера и структура поперечно–полосатого мышечного волокна. Струк-
турно-функциональной единицей скелетной мышцы является симпласт или мышечное волокно – клетка больших размеров, имеющая форму протяженного цилиндра с заостренными краями. Длина мышечной клетки чаще всего соответствует длине целой мышцы и достигает 14 см, а диаметр равен нескольким сотым долям миллиметра. Мышечное волокно окружено оболочкой – сарколеммой. Скелетные мышцы прикреплены в основном к костям. Их сокращение инициируется нервными импульсами и подчиняется сознательному контролю. Снаружи отдельные мышечные волокна окружены рыхлой соединительной тканью, которая содержит кровеносные, лимфатические сосуды и нервные волокна. Группы мышечных волокон образуют пучки, которые, в свою очередь, объединяются в целую мышцу, помещенную в п лотный чехол соединительной ткани, переходящей на концах мышцы в сухожилия, крепящиеся к кости. Базальная мембрана, окружающая каждое мышечное волокно, синтезируется с помощью мышечного волокна, эндомизиальных фибробластов и эндотелиальных клеток. Она состоит из сети коллагена типа IV, которая связана с другими белками. Отдельное мышечное волокно окружается эндомизиальными коллагеновыми волокнами, которые сливаются с более толстыми коллагеновыми пучками, образующими перимизий, окружающий фасции волокон.
Крупные сосуды, нервы и мышечные веретена расположены в перимизии. Перимизий в свою очередь сливается с эпимизием, который является частью фасциальной ткани, окружающей целую мышцу и группу мышц. Эпимизий состоит в основном из коллагена типа I, перимизий из коллагена типа I
и III, тогда как эндомизий содержит коллагены типа III, IV и V. Коллаген типа IV и V ассоциирован с базальными мембранами мышечных волокон. Коллагеновые структуры мышц важны для обеспечения их пассивных эластических свойств, т.е. резистентности к пассивному растяжению и восстановлению инициальной длины во время циклических преобразований.
Клетка окружена плазматической мембраной – сарколеммой, которая покрыта сетью коллагеновых волокон, придающих ей прочность и эластичность. Длина отдельных мышечных клеток может достигать 10 см (портняжная мышца) и даже 50 см, толщина до 0,1 мм. К мышечному волокну подходят окончания двигательных нервов и множество кровеносных сосудов.
Внутренность мышечного волокна заполнена саркоплазмой, представляющей собой вязкую жидкость, содержащую ядра, митохондрии, миоглобин
и нитевидные миофибриллы толщиной 1–3 мкм каждая и располагающихся от одного конца мышечного волокна к другому. Красный цвет саркоплазмы обусловлен присутствием в ней миоглобина – внутриклеточного дыхательного пигмента, благодаря которому создается запас кислорода. Миофибриллы находятся в окружении саркоплазматического ретикулума, который прини-

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 15 Поперечно-полосатая мышечная ткань. Нервная ткань
мает участие в процессах роста, развития и восстановления мышцы. Взаимосвязанные мембранные трубочки находятся в узком пространстве между миофибриллами, окружая их и располагаясь параллельно между ними.
Помимо многоядерности отличительной чертой мышечного волокна является наличие в цитоплазме тонких волоконец – миофибрилл, расположенных вдоль клетки и уложенных параллельно друг другу, состоящих из актиновых и миозиновых нитей. Число миофибрилл в волокне достигает двух тысяч. Структурными единицами миофибриллы являются саркомеры, которые располагаются вдоль мышечных волокон через каждые 2,3 мкм. Миофибриллы являются сократительными элементами клетки и обладают способностью уменьшать свою длину при поступлении нервного импульса, стягивая тем самым мышечное волокно. Под микроскопом видно, что миофибрилла имеет поперечную исчерченность – чередующиеся темные и светлые полосы. При сокращении миофибриллы светлые участки уменьшают свою длину и при полном сокращении исчезают вовсе. Для объяснения механизма сокращения миофибриллы около пятидесяти лет назад Хью Хаксли была разработана модель скользящих нитей, затем она нашла подтверждение в экспериментах и сейчас является общепринятой.
Кроме основных сократительных белков актина и миозина в построении саркомера и его работе участвует десмин и тропомиозин (рис. 3.15 ). Актин – белок, способный активировать гидролиз АТФ. Актин может существовать в виде мономера (G -актин, «глобулярный актин») или полимера (F -актин, «фибриллярный актин»). Молекула актина состоит из двух доменов.
Актин и миозин – основные сократительные белки. Каждый моно-
мер актина содержит одну молекулу прочно связанной АТР. Исторически домены называются большим и малым, хотя их размеры практически одинаковы. Между доменами существует глубокая щель. При полимеризации G -актина в F -актин ориентация всех мономеров одинакова, поэтому F -актин обладает полярностью. Волокна F -актина имеют два разноименно заряженных конца – (+) и (-), которые полимеризуются с различной скоростью. В мышечных клетках эти концы стабилизированы специальными белками. Полимеризованный актин внешне похож на две скрученные друг относительно друга нитки бус, где каждая бусина представляет собой мономер актина.
Элементарной структурной единицей толстых нитей саркомера является молекула миозина. В настоящее время описано более десяти различных видов молекул миозина. В состав молекулы миозина скелетных мышц входят 6 полипептидных цепей – две тяжелые цепи миозина и четыре легкие цепи миозина. Эти цепи прочно ассоциированы друг с другом (нековалентными связями) и образуют единый ансамбль, который и является молекулой миозина. Тяжелые цепи миозина имеют сильно асимметричную структуру.

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 15 Поперечно-полосатая мышечная ткань. Нервная ткань
Рис. 3.15. Тропониновый комплекс, обнаруженный в составе тонких филаментов мышечного сократительного аппарата, состоящий из трех белковых субъединиц: тропонина I, тропонина Т и тропонина С
У каждой тяжелой цепи есть длинный спиралевидный хвост и маленькая компактная грушевидная головка. Спирализованные хвосты тяжелых цепей миозина скручены между собой наподобие каната. Этот канат обладает довольно высокой жесткостью, и поэтому хвосты молекулы миозина образуют палочковидные структуры. В нескольких местах жесткая структура нарушена. В этих местах расположены шарнирные участки, обеспечивающие подвижность отдельных частей молекулы миозина.
С каждой головкой тяжелой цепи миозина связана одна регуляторная
и одна существенная легкая цепь миозина. Обе легкие цепи миозина влияют на способность миозина взаимодействовать с актином и участвуют в регуляции мышечного сокращения.
Палочкообразные хвосты могут слипаться друг с другом за счет электростатических взаимодействий. При этом молекулы миозина могут располагаться либо параллельно относительно друг друга, либо антипараллельно. Комбинация параллельной и антипараллельной упаковок приводит к формированию биполярных (двухполюсных) филаментов миозина.
Половина молекул миозина повернута своими головами в одну сторону, а вторая половина – в другую сторону. Биполярный миозиновый филамент располагается в центральной части саркомера.
Головки миозина могут дотягиваться до нитей актина и контактировать с ними. При замыкании таких контактов образуются поперечные мостики, которые генерируют тянущее усилие и обеспечивают скольжение нитей актина относительно миозина.
Сердечная поперечно-полосатая мыщечная ткань. Саркомер – уча-
сток мышечного волокна, ограниченный двумя Z -линиями. Они отделяют участки двух соседних саркомеров, которые содержат только нити актина
![]()
МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 15 Поперечно-полосатая мышечная ткань. Нервная ткань
и составляют вместе I-полосу (I-диск). На срезе саркомера участок, занятый М -линией и прилежащими зонами, в которых располагаются только миозиновые нити, носит название Н -полосы (светлой зоны), а участок, в котором располагаются нити миозин и частично актина, – А -полосы (А -диска).
Существенную роль в сокращении и расслаблении играет эндоплазматический ретикулум, который в мышечной ткани называется саркоплазматическим (рис. 3.16 ).
Именно в тяжелой фракции саркоплазматического ретикулума и были обнаружены Са -каналы.
Рис. 3.16. Саркоплазматический ретикулум
Саркоплазматический ретикулум – это внутриклеточная мембранная система взаимосвязанных уплощенных пузырьков и канальцев (цистерн), которая окружает саркомеры миофибрилл. В скелетных мышцах саркоплазматический ретикулум морфологически разделяется на два отдела: терминальные цистерны (ТЦ), контактирующие с трубочками Т -системы (ТТ) – впячиваниями плазматической мембраны (ПМ), и продолговатые трубочки, расположенные в центральной части саркомеров. С помощью электронной микроскопии было обнаружено, что мембраны терминальных цистерн саркоплазматического ретикулума непосредственно соединены с мембранами трубочек Т -системы посредством соединительных ножек (СН). Терминальные цистерны саркоплазматического ретикулума двух соседних саркомеров, связанные соединительными ножками с трубочкой Т -системы, образуют триаду. Фрагменты триад были найдены в тяжелой фракции саркоплазматического ретикулума.

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 15 Поперечно-полосатая мышечная ткань. Нервная ткань
Легкая фракция саркоплазматического ретикулума состоит из фрагментов продолговатых трубочек ретикулума и отличается от тяжелой фракции высоким содержанием белка Са -АТФазы.
Под влиянием потенциала действия, который распространяется по цитолемме и Т -трубочкам, ионы кальция высвобождаются, поступают к миофибриллам и инициируют сократительный акт, взаимодействуя с регуляторными белками. После чего актиновые и миозиновые миофиламенты получают возможность взаимодействовать друг с другом специализированными боковыми цепочками и перемещаться друг другу навстречу.
Сердечная поперечно-полосатая мышечная ткань развивается из симметрических участков висцерального листка спланхнотома в шейной части тела зародыша. Эти участки называются миоэпикардиальной пластинкой, большинство клеток которой дифференцируются в сердечные миоциты (кардиомиоциты), остальные клетки мезотелия эпикарда. В ходе гистогенеза дифференцируется несколько видов кардиомиоцитов: сократительные, проводящие, переходные (промежуточные), а также секреторные. В отличие от скелетной поперечно-полосатой мышечной ткани ядра располагаются в центральной части клетки. Большинство ядер полиплоидны. У полюсов ядра в цитоплазме находятся обычные органоиды. Агранулярная эндоплазматическая сеть хорошо развита. Она формирует субсарколемальные системы, прилежащие к Т -системам. Здесь же имеются включения гликогена и липидов. Митохондрии образуют цепочки вокруг специальных органелл – миофибрилл.
Миофибриллы построены из постоянно существующих упорядоченно расположенных нитей актина и миозина. Для закрепления миофибрилл служат особые структуры – телофрагмы и мезофрагмы. Телофрагмы представляют собой сети из белковых молекул, натянуты поперек клетки и прикреплены к цитолемме. На продольном срезе кардиомиоцита они выглядят линиями толщиной около 100 нм, получившими название Z -линии. Посередине саркомера располагается мезофрагма (М -линии на продольном срезе). От мезофрагмы в сторону телофрагмы отходят нити миозина. От телофрагмы навстречу им – нити актина. Они встречаются и на некотором расстоянии идут параллельно, причем каждый толстый (миозиновый) филамент сопровождется шестью тонкими (актиновыми) миофиламентами.
Клетки сократительных кардиомиоцитов имеют удлиненную форму и в конце соединяются друг с другом так, что цепочки кардиомиоцитов составляют функциональные волокна толщиной 10–20 мкм, а области контакта образуют вставочные диски. Их боковые поверхности покрыты базальной мембраной, в которую снаружи вплетаются тонкие ретикулярные и коллагеновые волокна.
Нервная ткань. Общая характеристика.Нервная ткань лежит в осно-
ве нервной системы организма – сложной пространственной структуры в виде единой сети с многочисленными связями как на уровне отдельной клетки,

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 15 Поперечно-полосатая мышечная ткань. Нервная ткань
так и клеточных ансамблей. Нервная система регулирует и координирует физиологические процессы отдельных клеток, тканей, органов, их систем и организма в целом, хранит информацию, интегрирует, перерабатывает сигналы, поступающие из внешней и внутренней среды.
Гистологические элементы нервной ткани (нейроны и глиоциты) и органов чувств развиваются из нескольких источников (рис. 3.17 ). В ходе нейруляции образуется нейэктодерма, формируются нервная трубка, нервный гребень и нейрогенные плакоды. В нейроонтогенезе происходит ряд морфогенетических процессов (например, гибель нейронов, направленный рост аксонов). Их совокупный эффект приводит к образованию нервной системы, функционирование которой conditio sine gua non определяют синапсы – специализированные межклеточные контакты между нейронами, а также между нейронами и исполнительными элементами (мышечными и секреторными).
Рис. 3.17. Нейрон: 1 – тело нервной клетки; 2 – аксон; 3 – дендриты
Нервная ткань состоит из нейронов, выполняющих специфическую функцию, и нейроглии, обеспечивающей существование и специфическую функцию клеток нервной ткани и осуществляющей опорную, трофическую разграничительную, секреторную и защитную функции.
Нейробласты – клетки с большим округлым ядром, плотным ядрышком и бледной цитоплазмой – дают начало всем нейронам ЦНС. Нейроны – классический пример клеток, относящихся к статической популяции. Ни при каких условиях они in vivo не способны к пролиферации и обновлению. Обонятельные нейроны (происходят из обонятельных плакод) эпителиальной выстилки носовых ходов – единственное известное исключение.
Глиобласты – предшественники макроглии (астроциты и олегодендроглиоциты – глиоциты). Все типы макроглии способны к пролиферации.
Генез клеток микроглии спорен. Согласно наиболее распространенной точке зрения, клетки микроглии относятся к системе мононуклеарных фагоцитов. Достаточно вероятным представляется их нейроэктодермальный генез.
В этом случае подразумевается гетерогенность популяции клеток микрогии.
В постнатальном онтогенезе не происходит образования новых нейро-
цитов, или другими словами, погибающие нейроны не восстанавливаются.

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 15 Поперечно-полосатая мышечная ткань. Нервная ткань
Но из этого не следует, что в нервной системе отсутствует регенегация. Она осуществляется за счет восстановления целостности поврежденных нейронов, роста их отростков, размножения глиальных и швановских клеток.
Нейроциты. Нейроны (термин предложил Вильгельм фон Вальдейер) – главные клеточные типы нервной ткани. Это возбудимые клетки осуществляют передачу электрических сигналов (между собой при помощи нейромедиаторов в синапсах) и обеспечивают способность мозга к переработке информации.
Нейроциты различных отделов нервной системы значительно отличаются друг от друга по функциональному значению и морфологическим особенностям. В зависимости от функции нейроны делятся на рецепторные (чувствительные, или афферентные), ассоциативные и эффекторные (эфферентные). Афферентные нейроциты генерируют нервый импульс под влиянием различных воздействий внешней или внутренней среды организма. Ассоциативные (вставочные) нервные клетки осуществляют взаимодействие между нейронами. Эффекторные нейроциты передают возбуждение на ткани рабочих органов, побуждая их к действию. Размеры нейронов значительно варьируются, так диаметр тела клетки колеблется от 4–6 мкм до 130 мкм. Вариабельна и специфична также форма клеток различных отделов нервной системы.
Характерной чертой для всех зрелых нейронов является наличие у них отростков, которые обеспечивают проведение нервного импульса. По функциональному значению отростки нейронов делятся на два вида. Отростки, выполняющие функцию отведения нервного импульса обычно от тела нейрона, называются аксонами или нейритами. Нейрит заканчивается концевым аппаратом либо на другом нейроне, либо на тканях рабочего органа. Дендриты, которые сильно ветвятся, приводят импульс к телу нейроцита. Количество и длина дендритов, характер их ветвления специфичны для различных типов нейроцитов. По количеству отростков нейроны делят:
на униполярные – с одним отростком;
псевдоуниполярные – с двумя отростками, которые имеют общий ствол отхождения от тела нервной клетки;
биполярные – с двумя отдельными отростками (аксоном и дендритом);
мультиполярные – с тремя и более отростками, при этом 1 аксон
и остальные дендриты.
Мультиполярные нейроны наиболее распространены у млекопитающих животных и человека.
Кроме отростков у нейроцитов различают перикарион (тело), в центре которого располагается обычно одно ядро. Форма ядра нейронов округлая содержащая, в основном эухроматин и 1–2 и более крупных,четких ядрышек, что соответствует высокой функциональной активности.
Двуядерные и многоядерные нейроциты встречаются очень редко. Исключение составляют нервные клетки некоторых ганглиев вегетативной

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 15 Поперечно-полосатая мышечная ткань. Нервная ткань
нервной системы (например, нейроны шейки матки и предстательной железы могут содержать до 15 ядер).
Наиболее развитыми органеллами цитоплазмы клетки является гранулярный и агранулярный эндоплазматический ретикулум (ЭР). Наличие развитого зернистого ЭР и его локализация в перикарионе доказано с помощью окраски нейроцита основными красителями по методу Ниссиля. При микроскопировании препаратов базофильные глыбки имеют пятнистый вид, и эту область цитоплазмы нейроцита называют тигроидом. В цитоплазме локализованы рибосомы, митохондрии, лизосомы, клеточный центр. Особую роль
в обеспечении специфической функциональной активности нейрона играет плазмолемма, которая способна проводить возбуждение, связанное с быстрым перемещением локальной деполяризации плазмолеммы по её дендритам к перикариону и аксону.
Под плазмолеммой перикариона располагается цитоскелет, состоящий из микротрубочек, промежуточных филаментов (нейрофиламентов) и микрофиламентов. Отростки нервных клеток также имеют субмемранную систему, элементы которой ориентированы паралельно в составе дендритов и нейритов, включая их тончайшие концевые ветвления. Микротрубочки – наиболее крупные элементы цитоскелета, их диаметр 24 нм. С ними связывают внутриклеточный, в том числе аксонный транспорт. От перикариона к отросткам перемещаются различные вещества и органеллы. Микротрубочки в перикарионе и дендритах не имеют направленной ориентации. В аксоне большинство микротрубочек ориентированы (+) – концом к терминали, а (–) – концом к перикариону. Характер ориентации микротрубочек имеет важное значение для распределения по отросткам различных органелл. К (+) – концу перемещаются митохондрии и секреторные пузырьки, а к (–) – концу – рибосомы, мультивезикулярные тельца, элементы аппарата Гольджи.
Способность синтезировать и секретировать биологически активные вещества свойственна всем нейроцитам. Однако существуют нейроциты, которые специализированы преимущественно для этой функции – секреторные нейроциты. Эти клетки имееют специфические морфологические признаки. Секреторные нейроны – крупные клетки, в цитоплазме и в аксонах находятся различной величины гранулы нейросекрета. Нейросекреты выбрасываются
в кровь и/или мозговую жидкость и играют роль нейрорегуляторов, обеспечивая взаимодействие нервной и гуморальной интеграции.
Нейроглия. Межклеточным веществом нервной ткани является ней-
роглия. Она имеет клеточное строение и выполняет в нервной ткани опорную, разграничительную, трофическую, секреторную и защитную функции. Клетки нейроглии составляют почти половину объема мозга. Все клетки нейроглии делят на два генетически различных вида: глиоциты (макроглия) и микроглию. Среди глиоцитов различают эпендимоциты, астроциты и олигодендроциты.
Эпендимоциты образуют плотный слой клеточных элементов, выстилающих спинномозговой канал и все желудочки мозга. В процессе гистоге-

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 15 Поперечно-полосатая мышечная ткань. Нервная ткань
неза нервной ткани эти глиальные клетки дифференцируются первыми из глиобластов нервной трубки и выполняют на этой стадии развития разграничительную и опорную функции. Вытянутые тела глиобластов на внутренней поверхности нервной трубки образуют слой эпителиоподобных клеток. На поверхности клеток, обращенной в полость канала нервной трубки, дифференцируются реснички. Мерцательные колебания ресничек способствуют движению спинномозговой жидкости. Базальные концы эпендимоцитов снабжены длинными отростками, которые, разветвляясь, пересекают всю нервную трубку, образуя её поддерживающий аппарат. Отростки, достигая внешней поверхности нервной трубки, принимают участие в образовании поверхностной глиальной пограничной мембраны, отделяющей вещество трубки от других тканей. Некоторые эпендимоциты выполняют секреторную функцию, выделяя различные активные вещества прямо в полость мозговых желудочков или кровь.
Эпендимоциты сосудистых сплетений желудочков мозга имеют кубическую форму.
На базальном полюсе образуются многочисленные и глубокие складки, цитоплазма содержит крупные митохондрии и включения. Есть сведения, что эпендимоциты участвуют в образовании цереброспинальной жидкости и регуляции её состава.
Астроциты – звездчатые клетки, их отростки отходят от тела клетки в разных направлениях, оплетают нейроны, сосуды, клетки эпендимы желудочков мозга, образуя расширения в виде концевой ножки. Различают волокнистые астроциты с длинными, слабо или совсем неветвящимися отростками. Они лежат преимущественно в белом веществе мозга. Протоплазматические астроциты с многочисленными короткими и ветвящимися отростками находятся в сером веществе мозга.
Функции астроцитов многочисленны:
в гистогенезе создают проводящие пути для миграции недифференцированных нейронов в коре мозжечка и для врастаний аксонов в зрительный нерв;
транспортируют метаболиты из капилляров мозга в нервную ткань, астроцитарные ножки почти полностью покрывают капилляры мозга;
участвуют в регуляции состава межклеточной жидкости, метаболизме возбуждающего и тормозного нейромедиаторов ЦНС;
изолируют рецептивные поверхности нейронов;
выделяют ряд веществ, способствующих росту аксонов. Олигодендроглиоциты составляют самую многочисленную группу кле-
ток нейроглии. В разных отделах нервной системы олидендроциты имеют различную форму. Они окружают тела нейронов в центральной и периферической нервной системе. Олигодендроциты являются миелинообразующими клетками ЦНС и имеют высокую плотность органелл. В белом веществе мозга олигодендроглиоциты расположены рядами между нервными волокнами,

МОДУЛЬ 3 ОСНОВЫ ГИСТОЛОГИИ
Лекция 15 Поперечно-полосатая мышечная ткань. Нервная ткань
и именно миелин придает белому веществу характерный цвет, отличающий его от серого вещества. Олигодендроглиоцины играют важную роль в образовании оболочек вокруг отростков (аксонов) клеток, и при этом они называются леммоцитами или швановскими клетками.
Микроглия представлена мелкими, неправильной формы клетками, имеющими многочисленные ветвящиеся отростки. В цитоплазме находится ядро с крупными глыбками хроматина, множество лизосом и плотные пластинчатые тельца. Клетки микролгии способны к амебовидному движению и в ответ на повреждение нервной ткани быстро размножаются и активизируются.
Отростки нервных клеток, обычно покрытые оболочками, называют нервными волокнами. В различных отделах нервной системы оболочки нервных волокон значительно отличаются друг от друга по своему строению. Все нервные волокна делятся на миелиновые и безмиелиновые.
Глава 5. ОСНОВНЫЕ ПОНЯТИЯ ОБЩЕЙ ГИСТОЛОГИИ
Глава 5. ОСНОВНЫЕ ПОНЯТИЯ ОБЩЕЙ ГИСТОЛОГИИ
Ткань - это возникшая в ходе эволюции частная система организма, состоящая из одного или нескольких дифферонов клеток и их производных, обладающая специфическими функциями благодаря кооперативной деятельности всех ее элементов.
5.1. ТКАНЬ КАК СИСТЕМА
Любая ткань - сложная система, элементы которой - клетки и их производные. Сами ткани тоже являются элементами морфофункциональных единиц, а последние выступают в роли элементов органов. Поскольку по отношению к системе высшего ранга (в нашем случае - организму) системы более низких рангов рассматриваются как частные, то и о тканях следует говорить как о частных системах.
В любой системе все элементы упорядочены в пространстве и функционируют согласованно друг с другом; система в целом обладает при этом свойствами, не присущими ни одному из ее элементов, взятому в отдельности. Соответственно и в каждой ткани ее строение и функции несводимы к простой сумме свойств отдельных входящих в нее клеток и их производных. Ведущими элементами тканевой системы являются клетки. Кроме клеток, различают клеточные производные (постклеточные структуры и симпла-сты) и межклеточное вещество (схема 5.1).
Среди клеточных структур целесообразно различать те, которые, будучи рассматриваемы и вне ткани, полностью обладают свойствами живого (например, способностью к размножению, регенерации при повреждениях и т. п.), и те, которые не обладают полнотой свойств живого. Постклеточные (послеклеточные) структуры относятся к последним.
Клеточные структуры, прежде всего, могут быть представлены индивидуально существующими клетками, каждая из которых имеет собственное ядро и собственную цитоплазму. Такие клетки могут быть либо одноядер-
Схема 5.1. Основные структурные элементы тканей
ными, либо многоядерными (если на каком-то этапе произошла нуклеото-мия без цитотомии). Если клетки по достижении какого-либо этапа развития сливаются друг с другом, то возникают симпласты. Примерами их могут служить симпластотрофобласт, остеокласты и симпластическая часть мышечного волокна скелетной мышечной ткани. Симпласты имеют совершенно иной принцип возникновения, нежели многоядерные клетки, так что эти понятия смешивать нецелесообразно.
Особо следует упомянуть случай, когда при делении клеток цитотомия остается незавершенной и отдельные из них остаются соединенными тонкими цитоплазматическими мостиками. Это - синцитий. Такая структура у млекопитающих встречается только в ходе развития мужских половых клеток, однако, поскольку эти клетки не относятся к соматическим, данную структуру не приходится причислять к тканевым.
Постклеточными структурами называют те производные клеток, которые утратили (частично или полностью) свойства, присущие клеткам как живым системам. Несмотря на это, постклеточные структуры выполняют важные физиологические функции, их нельзя расценивать просто как отмирающие или погибшие клетки. Среди постклеточных структур различают производные клеток в целом и производные их цитоплазмы. К первым относятся эритроциты большинства млекопитающих (форменные элементы крови, утратившие ядро на одном из этапов своего развития), роговые чешуйки эпидермиса, волосы, ногти. Примером вторых могут служить тромбоциты (производные цитоплазмы мегакариоцитов).
Межклеточное вещество - продукты синтеза в клетках. Его подразделяют на основное («аморфное», матрикс) и на волокна. Основное вещество может существовать в формах жидкости, золя, геля или быть минерализованным. Среди волокон различают обычно три вида: ретикулярные, коллагеновые и эластические.
Клетки всегда находятся во взаимодействии друг с другом и с межклеточным веществом. При этом формируются различные структурные объединения. Клетки могут лежать в межклеточном веществе на расстоянии друг от друга и взаимодействовать через него без непосредственных контактов (например, в рыхлой волокнистой соединительной ткани), либо соприкасаясь отростками (ретикулярная ткань) или образуя сплошные клеточные массы, или пласты (эпителий, эндотелий).
Дистантно клетки могут взаимодействовать с помощью химических соединений, которые клетки синтезируют и выделяют в процессе своей жизнедеятельности. Такие вещества служат не в качестве внешних секретов, как, например, слизь или пищевые ферменты, а выполняют регуля-торные функции, действуя на другие клетки, стимулируя или тормозя их активность. На этой основе формируется система положительных и отрицательных обратных связей, образуя управляющие контуры. Для реализации каждой из связей требуется некоторое время. Поэтому в тканях активность их жизнедеятельности не остается строго постоянной, а колеблется вокруг некоего среднего состояния. Такие регулярные колебания являются проявлением биологических ритмов на тканевом уровне.
Среди регуляторных веществ (иногда их называют биологически активными веществами) различают гормоны и интеркины. Гормоны поступают в кровь и способны действовать на значительных расстояниях от места их выработки. Интеркины действуют местно. К их числу относят вещества, угнетающие и стимулирующие клеточное размножение, определяющие направления дифференцировки клеток-предшественников, а также регулирующие запрограммированную клеточную гибель (апоптоз).
Таким образом, все межклеточные взаимодействия, как непосредственные, так и через межклеточное вещество, обеспечивают функционирование ткани как единой системы. Только на основе системного подхода возможно изучение тканей, понимание общей гистологии.
5.2. РАЗВИТИЕ ТКАНЕЙ (ЭМБРИОНАЛЬНЫЙ ГИСТОГЕНЕЗ)
В эмбриогенезе человека наблюдаются все процессы, характерные для позвоночных животных: оплодотворение, образование зиготы, дробление, гаструляция, формирование трех зародышевых листков, обособление комплекса эмбриональных зачатков тканей и органов, а также мезенхимы, заполняющей пространства между зародышевыми листками.
Геном зиготы не активен. По мере дробления в клетках - бластомерах - отдельные части генома активизируются, причем в разных бластомерах - разные. Этот путь развития генетически запрограммирован и обозначается как детерминация. В результате появляются стойкие различия их биохимических (а также и морфологических) свойств - дифференцировка. Одновременно дифференцировка сужает потенции дальнейшей активации
генома, которая возможна теперь за счет его оставшейся неактивированной части - происходит ограничение возможностей развития - комми-тирование.
По времени дифференцировка не всегда совпадает с детерминацией: детерминация в клетках может уже совершиться, а специфические функции и морфологические особенности проявятся позже. Подчеркнем, что все эти процессы совершаются на уровне генома, но без изменения набора генов как целого: гены не исчезают из клетки, хотя они могут быть и не активными. Такие изменения называют эпигеномными, или эпигенетическими.
Вопрос о том, насколько возможен возврат активной части генома вновь в неактивное состояние (дедифференцировка) в естественных условиях, остается неясным (это не исключает таких возможностей при генно-инженерных экспериментах).
Дифференцировка и коммитирование в эмбриогенезе появляются не сразу. Они совершаются последовательно: сначала преобразуются крупные участки генома, детерминирующие наиболее общие свойства клеток, а позднее - более частные свойства. В развивающемся организме дифферен-цировка сопровождается специфической организацией или размещением специализирующихся клеток, что выражается в установлении определенного плана строения в ходе онтогенеза - морфогенеза.
В результате дробления зародыш разделяется на внезародышевую и зародышевую части, причем становление тканей идет и в той, и в другой. В результате гаструляции в зародышевой части формируются гипобласт и эпибласт, а далее - образуются три зародышевых листка. В составе последних вследствие детерминации обособляются эмбриональные зачатки (еще не ткани). Их клетки обладают такой детерминацией и, в то же время, коммитированием, что в естественных условиях они не могут превратиться в клетки другого эмбрионального зачатка. Эмбриональные зачатки в свою очередь представлены стволовыми клетками - источниками дифферонов, формирующих ткани в эмбриональном гистогенезе (рис. 5.1). Межклеточного вещества зачатки не имеют.
В процессе образования трех зародышевых листков часть клеток мезодермы выселяется в промежутки между зародышевыми листками и формирует сетевидную структуру - мезенхиму, заполняющую пространство между зародышевыми листками. В последующем дифференцировка зародышевых листков и мезенхимы, приводящая к появлению эмбриональных зачатков тканей и органов, происходит неодновременно (гетерохронно), но взаимосвязанно (интегративно).
На понятии «мезенхима» следует остановиться особо. Содержание, которое вкладывают в него, весьма разнообразно. Часто ее определяют как эмбриональную соединительную ткань либо как эмбриональный зачаток. В последнем случае говорят о развитии из мезенхимы конкретных тканей, на основе чего даже делают выводы о родственности этих тканей. Мезенхиму считают источником развития клеток фибробластического ряда и клеток крови, эндотелиоцитов и гладких миоцитов, клеток мозгового вещества надпочечников. В частности, такая концепция долгое время «обосновывала» принадлежность эндотелия к соединительной ткани с отрицани-

Рис. 5.1. Локализация эмбриональных зачатков тканей и органов в теле зародыша (срез зародыша в стадии 12 сомитов, по А. А. Максимову, с изменениями): 1 - кожная эктодерма; 2 - нервная трубка; 3 - нейральный гребень; 4 - дерматом; 5 - миотом; 6 - склеротом; 7 - сегментная ножка; 8 - выстилка целома; 9 - аорта, выстланная эндотелием; 10 - клетки крови; 11 - кишечная трубка; 12 - хорда; 13 - полость целома; 14 - мигрирующие клетки, образующие мезенхиму
ем его тканевой специфичности. В некоторых учебниках анатомии до сих пор можно встретить классификацию мышц (как органов) на основании их развития либо из миотомов, либо из мезенхимы.
Признание мезенхимы в качестве эмбриональной соединительной ткани вряд ли состоятельно, хотя бы потому, что клетки ее еще не обладают одним из основных свойств ткани - специфической функцией. Они не синтезируют коллаген, эластин, гликозаминогликаны, как это свойственно фибробластам соединительной ткани, они не сокращаются, как миоциты, не обеспечивают двустороннего транспорта веществ, как эндотелиоциты. Морфологически они неотличимы друг от друга. Вряд ли можно считать мезенхиму и единым эмбриональным зачатком: в ходе развития зародыша клетки многих из них выселяются в нее, будучи уже соответственно детерминированными.
В составе мезенхимы совершается, в частности, миграция промиобластов и миобластов (выселившихся из сомитов), предшественников меланоцитов и клеток мозгового вещества надпочечников, клеток АПУД-серии (высе-
лившихся из сегментов нейрального гребня), клеток-предшественников эндотелия (скорее всего, выселившихся из спланхнотомов) и другие. Можно полагать, что, мигрируя и вступая друг с другом в контактные или химические взаимоотношения, клетки могут детализировать свою детерминацию.
Во всяком случае, считать мезенхиму единым эмбриональным зачатком не приходится. В рамках эпигеномных представлений ее надо рассматривать как гетерогенное образование. Клетки мезенхимы, хотя и сходны по морфологическим признакам, вовсе не безлики и не однолики в эпигеном-ном смысле. Поскольку клетки мезенхимы дают начало многим тканям, ее называют также плюриили полипотентным зачатком. Такое понимание противоречит представлению о зачатках как клеточных группировках, в которых клетки уже достигли значительной степени коммитированности. Признание мезенхимы единым зачатком означало бы отнесение к одному типу таких тканей, как скелетная, мышечная, кровь, железистый эпителий мозгового вещества надпочечников и многих других.
Как уже было отмечено, говорить о происхождении какой-либо ткани из зародышевого листка совершенно недостаточно для характеристики свойств и принадлежности к гистогенетическому типу. Столь же малозначаще и постулирование развития какой-либо ткани из мезенхимы. Судьба клеток мезенхимы по завершении их миграции - дифференциация в клетки конкретных тканей в составе конкретных органов. После этого мезенхимы как таковой не остается. Поэтому концепции о так называемом мезенхимном резерве неправомерны. В составе дефинитивных тканей, безусловно, могут оставаться либо стволовые клетки, либо клетки-предшественники, но это - клетки с уже детерминированными гистиотипическими свойствами.
Диффероны. Совокупность клеток, ведущих свое начало от общей пред-ковой формы, можно рассматривать как ветвящееся дерево последовательных процессов детерминации, сопровождающихся при этом коммитиро-ванием путей развития. От клеток, у которых эти процессы совершаются на уровне эмбриональных зачатков, можно проследить отдельные ветви, ведущие к различным конкретным дефинитивным (зрелым) клеточным видам. Такие исходные клетки называют стволовыми, а совокупность ветвей их потомков объединяют в диффероны. В составе дифферона происходят дальнейшая детерминация и коммитирование потенций развития стволовой клетки, в результате чего возникают так называемые клетки-предшественники. В каждой из таких ветвей, в свою очередь, возникают уже зрелые дифференцированные клетки, которые затем стареют и отмирают (рис. 5.2). Стволовые клетки и клетки-предшественники способны к размножению и в совокупности могут быть названы камбиальными.
Так, в системе крови от единой стволовой клетки всех форменных элементов (см. более подробно в главе 7 «Кровь» и «Кроветворение») возникают общая ветвь гранулоцитов и моноцитов, общая ветвь различных видов лимфоцитов, а также не ветвящаяся эритроидная линия (иногда такие ветви и линии тоже рассматривают как отдельные диффероны).
Хотя стволовые клетки детерминируются еще в составе эмбриональных зачатков, они могут сохраняться и в тканях взрослых организмов, но их

Рис. 5.2. Схема организации клеточного дифферона:
Классы клеток в диффероне: I - стволовые клетки; II - полипотентные клетки-предшественники; III - унипотентные клетки-предшественники; IV - созревающие клетки; V - зрелые клетки; выполняющие специфические функции; VI - стареющие и гибнущие клетки. В классах I-III происходит размножение клеток, это отображено на схеме двумя стрелками, отходящими от клетки вправо. Митотическая активность при этом нарастает. Клетки классов IV-VI не делятся (вправо отходит лишь одна стрелка).
СК - стволовые клетки; КПП - клетки-предшественники полипотентные; КПУ - клетки-предшественники унипотентные; КСо - клетки созревающие (уже не делящиеся, но еще не имеющие окончательных специфических функций); КЗр - зрелые клетки (обладающие специфическими функциями); КСт - стареющие клетки (утрачивающие полноту специфических функций).
Цифры после указания на класс клеток условно означают номер поколения в данном классе, следующие за ними буквы - свойства клеток. Обратите внимание, что дочерние клетки, возникшие в результате последовательных делений (классы I-III), имеют разную детерминацию, но сохраняют ее свойства в классах IV-VI. Толстая стрелка слева, направленная вниз, - сигнал для деления стволовой клетки, после того как одна из них вышла из популяции и вступила на путь дифференци-ровки
собственных предков уже не остается. Поэтому в организме нет таких клеточных форм, которые могли бы восполнить убыль стволовых, если она по какой-либо причине произошла, поэтому важнейшее свойство стволовых клеток - самоподдержание их популяции. Это означает, что в естественных условиях, если одна из стволовых клеток вступает на путь дифференциации, и, таким образом, общая их численность снижается на одну, восстановление популяции происходит только за счет деления аналогичной стволовой клетки из той же популяции. При этом она полностью сохраняет свои исходные свойства. В диффероне самоподдерживающуюся клеточную
популяцию выделяют в класс I. Наряду с этим определяющим признаком, стволовые клетки обладают и более частными, но существенными, с медицинской точки зрения, свойствами: стволовые клетки делятся очень редко, следовательно, они наиболее устойчивы к повреждающим воздействиям. Поэтому в случае чрезвычайных ситуаций они гибнут в последнюю очередь. Пока стволовые клетки сохраняются в организме, клеточная форма регенерации тканей возможна после устранения вредоносных воздействий. Если пораженными оказались и стволовые клетки, то клеточная форма регенерации не происходит.
В отличие от стволовых клеток, численность популяции клеток-предшественников может пополняться не только за счет деления клеток, себе подобных, но и за счет менее дифференцированных форм. Чем далее заходит дифференцировка, тем меньшую роль играет самоподдержание, поэтому пополнение популяции дефинитивных клеток происходит, в основном, за счет деления предшественников на промежуточных этапах развития, а стволовые клетки включаются в размножение только тогда, когда активности промежуточных предшественников для пополнения популяции недостаточно.
Клетки-предшественники (иногда их называют полустволовыми) составляют следующую часть гистогенетического древа. Они коммитированы и могут дифференцироваться, но не по всем возможным, а лишь по некоторым направлениям. Если таких путей несколько, клетки называют полипо-тентными (класс II), если же они способны дать начало лишь одному виду клеток - унипотентными (класс III). Пролиферативная активность клеток-предшественников выше, чем у стволовых, и именно они пополняют ткань новыми клеточными элементами.
На следующем этапе развития деления прекращаются, но морфологические и функциональные свойства клеток продолжают изменяться. Такие клетки называют созревающими и относят к классу IV. По достижению окончательной дифференцировки зрелые клетки (класс V) начинают активно функционировать. На последнем этапе их специфические функции угасают и клетки гибнут путем апоптоза (стареющие клетки, класс VI). Направление развития клеток в диффероне зависит от многих факторов: в первую очередь, от интеркинов микроокружения и от гормональных.
Соотношение клеток различной степени зрелости в дифферонах разных тканей организма неодинаково. Клетки различных дифферонов в процессе гистогенеза могут объединяться, причем количество дифферонов в каждом виде тканей может быть различным. Клетки дифферонов, входящих в ткань, участвуют в синтезе ее общего межклеточного вещества. Результатом гистоге-нетических процессов является формирование тканей с их специфическими функциями, не сводимыми к сумме свойств отдельных дифферонов.
Итак, под тканями целесообразно понимать частные системы организма, относящиеся к особому уровню его иерархической организации и включающие в качестве ведущих элементов клетки. Клетки тканей могут относиться к единому или к нескольким стволовым дифферонам. Клетки
одного из дифферонов могут преобладать и быть функционально ведущими. Все элементы ткани (клетки и их производные) равно необходимы для ее жизнедеятельности.
5.3. КЛАССИФИКАЦИИ ТКАНЕЙ
Существенное место среди вопросов общей гистологии занимают проблемы классификации тканей. В отличие от формальных классификаций, отталкивающихся от удобных для наблюдения признаков, естественные классификации призваны учитывать глубокие природные связи между объектами. Именно поэтому структура любой естественной классификации отражает реальную структуру природы.
Время от времени классификационные схемы меняются. Это означает, что в изучении природы сделан еще один шаг, и закономерности исследованы более полно и точно. Разносторонность подходов к характеристикам предметов классификации определяет и многомерность классификационных схем.
С позиции филогенеза предполагается, что в процессе эволюции как у беспозвоночных, так и позвоночных образуются четыре тканевые системы, или группы. Они обеспечивают основные функции организма: 1 - покровные, отграничивающие его от внешней среды и разграничивающие среды внутри организма; 2 - внутренней среды, поддерживающие динамическое постоянство состава организма; 3 - мышечные, отвечающие за движение; и 4 - нервные (или нейральные), координирующие восприятие сигналов внешней и внутренней среды, их анализ и обеспечивающие адекватные ответы на них.
Объяснение этому феномену дали А. А. Заварзин и Н. Г. Хлопин, которые заложили основы учения об эволюционной и онтогенетической детерминации тканей. Так, было выдвинуто положение о том, что ткани образуются в связи с основными функциями, обеспечивающими существование организма во внешней среде. Поэтому изменения тканей в филогенезе идут параллельными путями (теория параллелизмов А. А. Заварзина). При этом дивергентный путь эволюции организмов ведет к возникновению все большего разнообразия тканей (теория дивергентной эволюции тканей Н. Г. Хлопина). Из этого следует, что ткани в филогенезе развиваются и параллельными рядами, и дивергентно. Дивергентная дифференциация клеток в каждой из четырех тканевых систем в конечном итоге и привела к большому разнообразию видов тканей.
Позже выяснилось, что в ходе дивергентной эволюции конкретные ткани могут развиваться не только из одного, а из нескольких источников. Выделение основного из них, дающего начало ведущему клеточному типу в составе ткани, создает возможности для классификации тканей по генетическому признаку, единство же структуры и функции - по морфо-физиологическому. Большинство гистологов сейчас опираются именно на

Схема 5.2. Развитие эмбриональных зачатков и тканей:
Арабские цифры - эмбриональные зачатки; римские цифры - этапы развития зародыша и гистогенеза; А-Г - группы тканей.
В основании схемы (I уровень) лежит зигота. На II уровень поставлена морула - форма строения зародыша, которая возникает на этапе дробления. На III уровне отмечена бластоциста. В ней выделяются эмбриобласт и трофобласт (IV уровень). С этого времени развитие идет дивергентно. В эмбриобласте выделяются два листка - эпибласт и гипобласт, показанные на V уровне.
Возникновение и развитие половых клеток выделено особым стилем линии. Они остаются недетерминированными вплоть до взрослого состояния организма и, соответственно, не коммитируются. Поэтому если эмбриональные зачатки определять как совокупность клеток с соответствующей детерминацией и коммитированием, то понятие зачатка к совокупности первичных половых клеток неприменимо. На втором этапе гаструляции возникают три зародышевых листка (VI уровень). Именно в зародышевых листках в конце гаструляции и происходит детерминация (и соответствующее коммитирование) эмбриональных зачатков (VII уровень). Локализация зачатков в теле зародыша отмечена на VII уровне добавлением буквы «а». В энтодерме детерминируется энтеродермальный зачаток (1 - источник эпителиев кишечника и органов, связанных с ним).
В зародышевой эктодерме детерминируются эпидермальный и нейральный зачатки (3 и 4). Механизм детерминации прехордальной пластинки (2) до сих пор вызывает дискуссии, поэтому на схеме она отмечена как особая ветвь, возникающая при диф-ференцировке эпибласта, но не входящая в какой-либо определенный зародышевый листок.
В мезодерме детерминируются следующие зачатки: ангиобласт (5 - источник сосудистого эндотелия), сангвинальный (6 - источник форменных элементов крови), десмальный (7 - от греческого «десмос» - соединять, связывать, источник соединительных тканей и стромы гемопоэтических тканей), миосоматический (8 - источник поперечнополосатой скелетной мышечной ткани), целонефродермальный (9 - источник выстилки целома, эпителиев почек и половых органов, а также сердечной мышечной ткани). С мезодермой рассматривается и хорда, где детерминируется хордальный зачаток (10).
Клетки, мигрирующие и образующие мезенхиму (11), обозначены стрелками, выделенными цветом.
В соответствии с ведущими функциями тканей, последние представлены четырьмя основными морфофункциональными группами (VIII уровень схемы). В каждой группе присутствуют клетки, берущие начало из разных эмбриональных зачатков. Они обозначены соответствующими арабскими цифрами
сочетание морфофункциональной классификации А. А. Заварзина с генетической системой тканей Н. Г. Хлопина (однако из этого не следует, что удалось построить совершенную классификацию, которая была бы общепризнанной).
В настоящее время можно представить следующую схему классификации тканей (схема 5.2). На ней римскими цифрами показаны основные узлы, отражающие развитие зародыша от зиготы через уровень становления зародышевых листков и, далее, - эмбриональных зачатков. Заглавными буквами обозначены основные ткани, относящиеся к главным четырем морфо-функциональным группам. Эмбриональные зачатки обозначены арабскими
цифрами. Каждая группа может быть образована несколькими дифферона-ми, относящимися к разным гистогенетическим типам, однако существуют и монодифферонные ткани.
Очень часто при описании тканей среди прочих их функций выделяют так называемую «защитную», хотя это, по сути дела, отражает лишь сугубо утилитарный медицинский, но не общебиологический подход. В действительности же все функции тканей обеспечивают, прежде всего, нормальное динамическое равновесие всех систем организма в обычных постоянно меняющихся условиях существования. Лишь иногда воздействие факторов, нарушающих равновесие, переходит допустимые границы. В таких случаях обычные реакции, действительно, интенсифицируются и мобилизуются для восстановления нарушенного равновесия, и, как следствие, их качественные взаимоотношения меняются. Именно в подобных случаях на базе физиологических реакций возникают защитные. Они направлены на нейтрализацию и на ликвидацию агента, ставшего из нормального раздражителя угрожающим. Таким образом, понятие защиты целесообразно применять лишь в условиях патологии, применительно же к норме стоит говорить о поддержании равновесных соотношений. В норме нет факторов, с которыми надо бороться и от которых следует защищаться, в нормальных условиях ткани работают, будучи уравновешенными между собой и с окружающей средой.
В соответствии с морфофункциональным принципом, целесообразно в рамках группы выделять подгруппы, например группу тканей внутренней среды подразделить на подгруппы кровь и лимфу с кроветворными тканями, волокнистые соединительные ткани и скелетные ткани. В группе нейральных тканей в одну подгруппу целесообразно выделить собственно нервную ткань (совокупность нейронов как систему, непосредственно обусловливающую ее функции) и глию (как совокупность тканей, непосредственно «обслуживающих» нейроны), а также микроглию. В группе мышечных тканей выделяют подгруппы гладких и поперечнополосатых (неисчерченных и исчерченных).
5.4. РЕГЕНЕРАЦИЯ ТКАНЕЙ
Знание основ эмбрионального гистогенеза необходимо для понимания теории регенерации, т. е. восстановления структуры биологического объекта после утраты части ее элементов. Соответственно уровням организации живого различают внутриклеточную, клеточную, тканевую, органную формы регенерации. Предметом общей гистологии является регенерация на тканевом уровне. У разных тканей возможности регенерации неодинаковы. Различают физиологическую и репаративную регенерацию. Физиологическая регенерация генетически запрограммирована. Репаративная регенерация происходит после случайной гибели клеток, например, в результате интоксикации (в том числе и алкогольной), воздействий постоянного природного радиационного фона, космических лучей на организм.
Таблица 5.1. Регенерационные возможности тканей

При физиологической регенерации популяция клеток обновляется постоянно. Дифференцированные зрелые клетки имеют ограниченный срок жизни и, выполнив свои функции, гибнут путем апоптоза. Убыль популяции клеток восполняется за счет деления клеток-предшественников, а последних - за счет стволовых клеток. Такие ткани называют обновляющимися. Примерами таких тканей (среди многих других) могут служить многослойный кожный эпителий и кровь.
В некоторых тканях активное размножение клеток идет до тех пор, пока не закончился рост организма. Далее физиологической регенерации в них не происходит, хотя и после завершения роста в них остаются малодиффе-ренцированные клетки. В ответ на случайную гибель специализированных клеток возникает размножение малодифференцированных клеток, и популяция восстанавливается. После восстановления клеточной популяции размножение клеток снова угасает. Такие ткани относят к растущим. Отдельными примерами их могут служить эндотелий сосудов, нейроглия, эпителий печени.
Есть и такие ткани, в которых после окончания роста размножение клеток не наблюдается. В этих случаях ни физиологическая, ни репаративная регенерация невозможна. Такие ткани называют стационарными. Примерами могут служить сердечная мышечная ткань и собственно нервная ткань (совокупность нейронов). У взрослого человека в таких тканях регенерация происходит лишь на внутриклеточном уровне.
Изложенное кратко иллюстрирует табл. 5.1.
Контрольные вопросы
1. Перечислите основные структурные элементы тканей.
2. Охарактеризуйте понятия зародышевый листок, эмбриональный зачаток, дифферон.
3. Дайте определение ткани с позиции клеточно-дифферонной организации.
4. Назовите формы регенерации тканей.
Гистология, эмбриология, цитология: учебник / Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др.. - 6-е изд., перераб. и доп. - 2012. - 800 с. : ил.
Слайд 2
ОПРЕДЕЛЕНИЕ ТКАНИ
ТКАНЬ - это филогенетически сложившаяся система клеток и неклеточных структур, имеющая общее происхождение, строение и выполняющая определенные функции. Основой и родоначальником всех тканей является клетка. Именно те клетки, которые лежат в истоке ткани и поддерживают ее существование на протяжении всей жизни организма, называют стволовыми.
Слайд 3
ВИДЫ ТКАНЕЙ
В организме человека различают четыре группы тканей: эпителиальные ткани, ткани внутренней среды: кровь и соединительные ткани, мышечные ткани, нервную ткань
Слайд 4
ЭПИТЕЛИАЛЬНАЯ ТКАНЬ
Эпителиальная ткань - «эпи» - сверху, «телио» - выстилаю. Название больше подходит к покровному эпителию, занимающему пограничное положение. Также различают железистый эпителий, в составе желез и сенсорный эпителий - рецепторные клетки органов чувств. Наибольшего внимания заслуживает покровный эпителий. Покровный эпителий - это пласт тесно сомкнутых клеток, лежащих на базальной мембране. Между собой клетки соединены десмосомами, а с базальной мембраной - полудесмосомами.
Слайд 5
ПОКРОВНЫЙ ЭПИТЕЛИЙ
Покровный эпителий всегда отвечает определенным закономерностям строения и функции: лежит на границе с внешней и внутренней средой - пограничная ткань, между клетками почти отсутствует межклеточное вещество, пласт клеток лежит на базальной мембране и поэтому клетки эпителия имеют полярность - апикальную и базальные части. Ткань растет только от базальной мембраны.
Слайд 6
не содержит кровеносных сосудов, питание клеток происходит диффузно, сквозь базальную мембрану; эпителий содержит множество чувствительных нервных окончаний; под базальной мембраной лежит рыхлая соединительная ткань с сетью кровеносных капилляров.
Слайд 7
ФУНКЦИИ И СВОЙСТВА ПОКРОВНОГО ЭПИТЕЛИЯ
защитная (механическая, физическая защиты); барьерная (непроницаем для веществ, разграничивают ткани); транспортная; всасывание; секреторная; эксекреторная (метаболиты); сенсорная (чувств.окончания); высокая регенерация (клеточная и внутриклеточная).
Слайд 8
ФИЛОГЕНЕТИЧЕСКАЯ КЛАССИФИКАЦИЯ ЭПИТЕЛИЯ
Эктодермальный (эпидермальный) - преимущественно многослойный (характерна защитная функция); Эндодермальный - однослойный (характерно всасывание); Целонефродермальный - мочевыводящие пути и целомический мешок; однослойный (барьерная и эксекреторная); Эпендимоглиальный - выстилает желудочки мозга и спинномозговой канал - однослойный (барьерная). Эндотелиальный (ангиодермальный) - принадлежность однослойной выстилки сосудов к эпителию оспаривается многими авторами.
Слайд 9
МОРФОФУНКЦИОНАЛЬНАЯ КЛАССИФИКАЦИЯ ЭПИТЕЛИЯ
многослойный (ороговевающий и неороговевающий) плоский кубический переходный однослойный (однорядный и многорядныи) плоский кубический призматический
Слайд 10
ОДНОСЛОЙНЫЙ ЭПИТЕЛИЙ
Однослойный эпителий - все клетки лежат на базальной мембране. Плоский эпителий эпителий - в канальцах почек, альвеолах легких - контактирует с внешней средой; эндотелий - не имеет контакта с внешней средой, выстилает стенки сосудов; мезотелий - выстилает внутренние полости брюшины, плевры и перикарда.
Слайд 11
КУБИЧЕСКИЙ ЭПИТЕЛИЙ
канальцы почек; протоки желез; всасывание, барьерная функция; энтодерма и мезодерма.
Слайд 12
ОДНОСЛОЙНЫЙ КУБИЧЕСКИЙ ЭПИТЕЛИЙ
Слайд 13
Слайд 14
ОДНОСЛОЙНЫЙ ПРИЗМАТИЧЕСКИЙ ОДНОРЯДНЫЙ ЭПИТЕЛИЙ
Слайд 15
ЦИЛЛИНДРИЧЕСКИЙ И ПРИЗМАТИЧЕСКИЙ ЭПИТЕЛИЙ
Цилиндрический и призматический однорядный простой - верхушка клеток обычная, функция барьерная; железистый - клетки вырабатывают слизь для защиты - желудок;
Слайд 16
ОДНОСЛОЙНЫЙПРИЗМАТИЧЕСКИЙ МНОГОРЯДНЫЙ РЕСНИТЧАТЫЙ (МЕРЦАТЕЛЬНЫЙ) ЭПИТЕЛИЙ
Слайд 17
МНОГОСЛОЙНЫЙ ЭПИТЕЛИЙ
Многослойный эпителий Плоский ороговевающий или эпидермис, состоит из 5 слоев: базальный; шиповатый; зернистый; блестящий; роговой
Слайд 18
МНОГОСЛОЙНЫЙ ПЛОСКИЙ ОРОГОВЕВАЮЩИЙ ЭПИТЕЛИЙ (ЭПИДЕРМИС)
Слайд 19
ПЛОСКИЙ НЕОРОГОВЕВАЮЩИЙ ЭПИТЕЛИЙ
Плоский неороговевающий выстилает: полость рта; глотки; пищевода; анальный отдел прямой кишки; роговицу глаза; влагалище.
Слайд 20
МЕЗЕНХИМА
Источником развития тканей внутренней среды в организме является мезенхима. Условно ее можно представить соединительной тканью эмбриона, которая заполняет пространство между зародышевыми листками. Она состоит из мелких, отростчатых клеток, которые, соединяясь, образуют сетевидную структуру. Значительную часть между клетками занимает межклеточное вещество. В процессе эмбриогенеза вся мезенхима преобразуется в различные ткани зрелого организма, поскольку сама она также развивается из различных источников.
Слайд 21
КЛАССИФИКАЦИЯ
Ткани внутренней среды (система крови и соединительные ткани). 1.Система крови А)кровь и лимфа Б) кроветворная (миелоидная и лимфоидная) 2.Соединительные ткани А) собственно соединительная ткань (волокнистая) Б) соединительная ткань со специальными свойствами 1. жировая 2. ретикулярная 3. слизистая 4. пигментная 5. скелетного типа (костная и хрящевая)
Слайд 22
КРОВЬ
Как любая ткань, кровь состоит из клеток - форменных элементов и межклеточного вещества - плазмы. Отличается кровь от других тканей по своим физическим свойствам, она жидкая и поэтому подчиняется еще и физическим законам, как все жидкости
Слайд 23
МАЗОК КРОВИ ЧЕЛОВЕКА. Окраска по Романовскому-Гимзе.
Слайд 24
МАЗОК КРОВИ ЧЕЛОВЕКА.
1 - эритроциты; 2 - лимфоциты (малый и большой); 3 и 4 моноцит; 4 - нейтрофильные гра-нулоциты (лейкоциты); 5 - эозинофильный гранулоцит (лейкоцит); 6 - базофильный гранулоцит (лейкоцит); 7-кровяные пластинки (тромбоциты).
Слайд 25
ФУНКЦИИ И СВОЙСТВА КРОВИ
Основное свойство крови - реактивность. Внешнее воздействие или изменения внутренней среды организма меняет нормальное соотношение компонентов крови. дыхательная трофическая регуляторная (гомеостатическая) защитная (иммунная) свертывающаяся транспортная экскреторная
Слайд 26
ПЛАЗМА
Плазма - представляет собой жидкую среду, в которой взвешены системы белков, углеводы и липиды, находятся неорганические соединения и ионы. Состав плазмы - 92% воды и сухой остаток (сухая плазма)
Слайд 27
БЕЛКИ ПЛАЗМЫ
Альбумины - обеспечивает онкотическое давление, строительный материал для клеток и тканей, участвует в переносе питательных веществ. Глобуллины - это защитные, иммунные белки - антитела. Фибриноген - участвует в свертывании крови. и Протромбин - глобулин - вырабатывает иммунные клетки, другие белки крови синтезируют клетки печени; из липидов - наиболее важен холестерин; среди углеводов - часто определяют глюкозу, сахар крови - в норме 6,6 ммоль\л.
Слайд 28
СОСТАВ КРОВИ
Плазма крови без свертывающих белков называется сыворотка. Соотношение форменных элементов и плазмы называется гематокрит, который колеблется в пределах 35-50%. Процентное соотношение форменных элементов называется гемограмма. Повышенное количество форменных элементов крови называется -цитоз, реже - филия, повышение количества компонентов плазмы - гипер. Понижение количества форменных элементов крови называется - пения, а понижение количества компонентов плазмы - гипо. Снижение количества всех форменных элементов называется малокровие - анемия.
Классификация болезней человека проводится по характеру течения: острые хронические болезни по уровню, на котором в организме выявляются специфические патологические изменения при болезни: молекулярные хромосомные клеточные тканевые органные заболевания всего организма
Слайд 32
Общепринятая классификация болезней:
Внутренние болезни (терапия) - это заболевания, главным методом лечения которых являются лекарства Хирургические болезни (хирургия) - это заболевания, главным методом лечения которых является операция Злокачественные заболевания (онкология) - это заболевания, в основе которых лежит неконтролируемый процесс размножения одного из видов клеток
Слайд 33
Классификация болезней
Наследственные болезни - это заболевания, причиной которых является дефект гена Болезни органов, участвующих в вынашивании беременности и родах (гинекология) Кожные болезни - это болезни, ключевым клиническим проявлением которых является поражение кожи Глазные болезни - это болезни, ключевым клиническим проявлением которых являются поражения глаза
Посмотреть все слайды