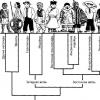
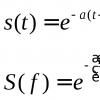Какие выделяют три типа мембранных белков. Организация и функции мембранных белков. Структура и функции клеточной мембраны
Как правило именно белки ответственны за функциональную активность мембран. К ним относятся разнообразные ферменты транспортные белки рецепторы каналы поры и. До этого считалось что мембранные белки имеют исключительно β складчатую структуру вторичная структура белка но данные работы показали что мембраны содержат большое количество α спиралей. Дальнейшие исследования показали что мембранные белки могут глубоко проникать в липидный бислой или даже пронизывать его и их стабилизация осуществляется за счёт гидрофобных...
Поделитесь работой в социальных сетях
Если эта работа Вам не подошла внизу страницы есть список похожих работ. Так же Вы можете воспользоваться кнопкой поиск
Лекция 5
Строение и функции мембранных белков
Клеточные мембраны содержат белка от 20 до 80% (по весу). Как правило, именно белки ответственны за функциональную активность мембран. К ним относятся разнообразные ферменты, транспортные белки, рецепторы, каналы, поры и. т.д., которые обеспечивают уникальность функций каждой мембраны. Первые успехи в изучении мембранных белков были достигнуты тогда, когда биохимики научились использовать детергенты для выделения мембранных белков в функционально активной форме. Это были работы по изучению ферментных комплексов внутренней мембраны митохондрий. До этого считалось, что мембранные белки имеют исключительно β складчатую структуру (вторичная структура белка), но данные работы показали, что мембраны содержат большое количество α спиралей. Значительно реже встречается β спираль, которой, однако, придают важное биологическое значение. Дело в том, что на участках, окружённых липидами, β спираль представляет собой полый цилиндр, в наружной стенке которого сосредоточены неполярные (гидрофобные) аминокислотные остатки, а во внутренней гидрофильные. Такой цилиндр мог бы образовать в мембране канал, через который свободно проходят ионы и водорастворимые вещества. Дальнейшие исследования показали, что мембранные белки могут глубоко проникать в липидный бислой или даже пронизывать его и их стабилизация осуществляется за счёт гидрофобных взаимодействий. Существует, как минимум, четыре вида расположения белков в мембранах: Первый вид трансмембранный, когда белок пронизывает всю мембрану, а гидрофобный участок белка имеет α конфигурацию. Похожее расположение в мембране имеет молекула бактериородопсина из Halobacterium halobium его α спирали последовательно пересекают бислой; Второй вид связывание с помощью гидрофобного якоря, когда у белка есть короткий участок, состоящего из гидрофобных остатков аминокислот вблизи карбоксильного конца. Это, так называемый, гидрофобный якорь, который можно удалить с помощью протеолиза, а высвобождённый белок становится водорастворимым. Такое расположение в мембране присуще многим цитохромам. Третий вид связывание с поверхностью бислоя, когда взаимодействие белков имеет в первую очередь электростатическую природу или гидрофобную природу. Данный тип взаимодействия может использоваться как дополнение к другим взаимодействиям, например, к трансмембранному заякориванию. Четвёртый тип- связывание с белками, погружёнными в бислой, это когда некоторые белки связываются с белками, которые располагаются внутри липидного бислоя. Например, F 1 - часть Н + - АТФазы, которая связывается с F 0 частью, погружённой в мембрану, а также некоторые белки цитоскелета.
В основе современных представлений о структуре мембранных белков лежит идея о том, что их полипептидная цепь уложена так, чтобы образовалась неполярная, гидрофобная поверхность, контактирующая с неполярной областью липидного бислоя. Полярные домены белковой молекулы могут взаимодействовать с полярными головками липидов на поверхности бислоя. Многие белки являются трансмембранными и пронизывают бислой. Некоторые белки, по видимому, связаны с мембраной лишь за счёт их взаимодействия с другими белками.
Многие мембранные белки обычно связываются с мембраной с помощью нековалентных взаимодействий. Однако есть белки, которые связаны с липидами ковалентно. Многие белки плазматических мембран относятся к классу гликопротеинов. Углеводные остатки этих белков всегда находятся с наружной стороны плазматической мембраны.
Обычно мембранные белки подразделяют на наружные (периферические) и внутренние (интегральные). При этом критерием служит степень жёсткости обработки, необходимой для извлечения этих белков из мембраны. Периферические белки высвобождаются при промывании мембран буферными растворами с низкой ионной силой, низким или, наоборот, с высоким значением рН и в присутствии хелатирующих агентов (например, ЭДТА), связывающих двухвалентные катионы. Часто бывает, что очень трудно отличить периферические мембранные белки от белков, связавшихся с мембраной в процессе выделения.
Для высвобождения интегральных мембранных белков необходимо использовать детергенты или даже органические растворители.
Многие мембранные белки эукариот и прокариот ковалентно связаны с липидами, которые присоединяются к полипептиду после трансляции.
Мембранные белки, ковалентно связанные с липидами
(А) Белки, к которым присоединена миристиновая кислота
(Б) Белки, к которым присоединена пальмитиновая кислота
(В) Белки с гликозилфосфатидилинозитольным якорем
4. Адгезивная молекула нервных клеток |
В некоторых случаях эти липиды играют роль гидрофобного якоря, с помощью которого белок прикрепляется к мембране. В других случаях липиды, вероятно, выполняют функцию помощника при миграции белка в соответствующую область клетки или (как в случае белков оболочки вирусов) в слиянии мембран.
У прокариот наиболее полно охарактеризован белок липопротеин Брауна основной липопротеин наружной мембраны E . coli . Зрелая форма этого белка содержит ацилглицерол, который связан тиоэфирной связью с N концевым цистеином. Кроме того, N концевая аминокислота связана с жирной кислотой амидной связью. Мембраносвязанная форма пенициллазы прикрепляется к цитоплазматической мембране с помощью N концевого ацилглицерола аналогично липопротеинам мембраны.
Мембранные белки эукариот ковалентно связанные с липидами, как показано в таблице, их можно разделить на три класса. Белки первых двух классов, по видимому, локализованы в основном на цитоплазматической поверхности плазматической мембраны, а белки третьего класса на наружной поверхности.
Существуют мембранные белки, которые ковалентно связаны с углеводами. К ним относятся поверхностные белки клеток в основном, выполняющих функции транспорта и рецепции. До сих пор неясно, в чём тут дело. Возможно, это связано с тем, что белки нужно сортировать при направлении их к плазматической мембране. Сахарные остатки могут защищать белок от протеолиза или участвовать в узнавании или адгезии. Поэтому сахарные остатки в мембранных гликопротеинах локализованы исключительно на наружной стороне мембраны.
Можно выделить два основных класса олигосахаридных структур мембранных гликопротеинов: 1) N гликозидные олигосахариды, связанные с белками через амидную группу аспаргина; 2) О-гликозидные олигосахариды, связанные через гидроксильные группы серина и треонина. Данный класс олигосахаридов состоит из трёх подклассов.
- Простой или обогащённый маннозой комплекс, в котором олигосахарид содержит маннозу и N ацетилглюкозамин.
- Нормальный комплекс, в котором обогащённый маннозой кор имеет дополнительные боковые ветви, содержащие другие сахаридные остатки, например сиаловую кислоту.
- Большой комплекс, который связан с анионным переносчиком мембраны эритроцитов
Большинство олигосахаридов мембранных гликопротеинов принадлежат к подклассу 1 или2.
Мембранные белки бактерий
Как уже отмечалось выше, белки в цитоплазматической мембране составляют около 50% её поверхности. Примерно 10% мембраны образовано прочно связанными белковолипидными комплексами. Молекула любого встроенного в мембрану белка окружена 45 130 и более липидными молекулами. Около половины свободных липидов связано с периферическими белками мембраны.
Белковый состав цитоплазматической мембраны бактерий более разнообразен, чем липидный. Так, в цитоплазматической мембране E . coli K 12 обнаружено около 120 различных белков. В зависимости от ориентации в мембране и характера связи с липидным бислоем, как уже отмечалось выше, белки делят на интегральные и периферические. К периферическим белкам бактерий можно отнести ряд ферментов таких как, НАДН дегидрогеназа, малатдегидрогеназа и др., а также некоторые белки, которые входят в состав АТФазного комплекса. Этот комплекс представляет собой группу определённым образом расположенных белковых субъединиц, контактирующих с цитоплазмой, периплазматическим пространством и образующих в мембране канал, через который осуществляется переход протона. Участок комплекса, обозначаемый F 1 , погружён в цитоплазму, а и с компоненты участка F 0 гидрофобными сторонами молекул погружены в мембрану. Субъединица b частично погружена в мембрану своей гидрофобной частью и осуществляет связь мембранной и цитоплазматической частей ферментного комплекса, а также связь синтеза АТФ в участке F 1 с протонным потенциалом в мембране. Субъединицы а, b и с обеспечивают протонный канал. Другие компоненты комплекса обеспечивают его структурную и функциональную целостность.
К интегральным белкам E . coli , которые для проявления энзиматической активности необходимы липиды, можно отнести сукцинатдегидрогеназу, цитохром b . Очень интересными свойствами обладает антибиотики грамицидин А, аламетицин, амфотерицин и нистацин. Они при взаимодействии с мембраной бактерий становятся интегральными белками (антибиотики являются полипептидами и макроциклами).
Грамицидин А это гидрофобный пептид, состоящий из 15 L - D -аминокислот. При встраивании в мембрану он образует каналы, которые пропускают одновалентные катионы. Этот канал, который образует грамицидин А охарактеризован наиболее полно. Канал образован двумя молекулами грамицидина А. В результате чередования L - и D - аминокислот образуется спираль, в которой боковые цепи располагаются снаружи, а карбоксильные группы остова внутри канала. Этот тип спирали, не встречается больше ни в каких белках и образуется из за необычного чередования стереоизомеров аминокислот в грамицидине А. Грамицидиновый канал, как уже отмечалось выше, катионселективен. Небольшие неорганические и органические катионы проходят через него, в тоже время проницаемость по Cl - равна нулю.
Аламетицин это пептидный антибиотик из 20 аминокислотных остатков, способный образовывать в мембране электовозбудимые каналы. Аминокислотная последовательность аламетицина включает необычные остатки α аминомасляную кислоту и L фенилаланин. При связывании с мембраной в отличие от грамицидина А он образует пору. Она намного по размеру меньше, чем канал, который образует грамицидин А. Прежде всего это связано с тем, что пространство вокруг α спирали слишком мало, чтобы через него мог пройти ион.
Марколидные антибиотки, такие как, нистатин и амфотерицин связываются с холестерином и образуют каналы. Каналы образуют 8 10 молекул этих полиеновых антибиотиков, через которые, правда, с невысокими скоростями проникают ионы.
Другие похожие работы, которые могут вас заинтересовать.вшм> |
|||
| 21572. | СТРОЕНИЕ И ФУНКЦИИ БЕЛКОВ | 227.74 KB | |
| Содержание белков в организме человека выше чем содержание липидов углеводов. Преобладание в тканях белков по сравнению с другими веществами выявляется при расчёте содержания белков на сухую массу тканей. Содержание белков в различных тканях колеблется в определённом интервале. | |||
| 17723. | Мозжечек, строение и функции | 22.22 KB | |
| 3 Общее строение головного мозга. В нервной системе выделяют также центральную часть ЦНС которая представлена головным и спинным мозгом и периферическую часть в которую входят нервы нервные клетки нервные узлы ганглии и сплетения топографически лежащие вне спинного и головного мозга. Объектом исследования является анатомия головного мозга. Данная цель предмет и объект подразумевают постановку и решение следующих задач: описать общий план строения головного мозга изучить анатомическое строение мозжечка выделить... | |||
| 5955. | Органы растений: их функции, строение и метаморфозы. | 16.94 KB | |
| Органы цветка являются видоизмененными листьями: покровные листья формируют чашелистики и лепестки а спорообразующие листья дают начало тычинкам и пестикам. Побег включает: а стебель б листья в вегетативные почки г цветки д плоды. Стеблем называется вегетативный орган растения который выполняет многочисленные функции: несёт листья или тяжелую крону из ветвей и листьев; связывает корни и листья; на нем образуются цветки; по нему передвигается вода с минеральными веществами и органическими соединениями; молодые стебли... | |||
| 5067. | Гладкие мышцы. Строение, функции, механизм сокращения | 134.79 KB | |
| Мышцы или мускулы от лат. Мышцы позволяют двигать частями тела и выражать в действиях мысли и чувства. Гладкие мышцы являются составной частью некоторых внутренних органов и участвуют в обеспечении функции выполняемые этими органами. | |||
| 6233. | Строение и функции ядра. Морфология и химический состав ядра | 10.22 KB | |
| От цитоплазмы ядра обычно отделяются четкой границей. Бактерии и синезеленые водоросли не имеют сформированного ядра: их ядро лишено ядрышка не отделено от цитоплазмы отчетливо выраженной ядерной мембраной и носит название нуклеоид. Форма ядра. | |||
| 9495. | Классификация, характеристика ассортимента пушно-мехового сырья и пушно-мехового полуфабриката, строение пушно-меховой шкуры, строение волоса и разновидность его форм, технология изготовления пушнины | 1.05 MB | |
| Меховые пластины полосы определенной формы сшитые из подобранных выделанных шкурок и предназначенные для раскроя на детали меховых изделий. К зимним видам пушного сырья относятся шкурки и шкуры пушных зверей добыча которых производится преимущественно в зимнее время когда качество шкурок особенно высоко. СТРОЕНИЕ И ХИМИЧЕСКИЙ СОСТАВ ШКУРОК ПУШНОМЕХОВОГО и овчинношубного СЫРЬЯ ПОНЯТИЕ О ТОПОГРАФИИ ШКУРКИ Шкуркой называют наружный покров животного отделенный от его тушки и состоящий из кожной ткани и волосяного покрова. У... | |||
| 8011. | Свойства мембранных липидов | 10.13 KB | |
| Некоторые липиды способствуют стабилизации сильно искривлённых участков мембраны образованию контакта между мембранами или связыванию определённых белков поскольку форма этих молекул благоприятствует нужной упаковке бислоя на соответствующих участках мембраны. Под жидкостным состоянием понимают способность фосфолипидных молекул к вращению и латеральному перемещению в соответствующем лепестке мембраны. Они вытянуты и ориентированы перпендикулярно плоскости мембраны. В состоянии жидкого кристалла молекулы жирных кислот подвижны но... | |||
| 8014. | Химический состав мембранных липидов | 10.81 KB | |
| Прежде всего это связано с множеством функций которые выполняют липиды в мембранах. Фосфатидная кислота в свободном виде содержится в мембранах бактерий в небольшом количестве обычно же к ней присоединены остатки спиртов аминокислот и др. Эти липиды являются сложными эфирами жирных кислот и глицерола и широко представлены во многих мембранах эукариотических и прокариотических клеток за исключением архебактерий. Они содержатся в большом количестве во внутренней мембране митохондрий в мембране хлоропластов и в некоторых бактериальных... | |||
| 21479. | ОБМЕН БЕЛКОВ | 150.03 KB | |
| Различают три вида азотистого баланса: азотистое равновесие положительный азотистый баланс отрицательный азотистый баланс При положительном азотистом балансе поступление азота преобладает над его выделением. При заболевании почек возможен ложный положительный азотистый баланс при котором происходит задержка в организме конечных продуктов азотистого обмена. При отрицательном азотистом балансе преобладает выделение азота над его поступлением. Это состояние возможно при таких заболеваниях как туберкулез ревматизм онкологические... | |||
| 15073. | Рассмотрение мембранных (ионоселективных) электродов с различного рода мембранами | 127.48 KB | |
| Для этого существуют разнообразные ионоселективные электроды главной особенностью которых является так называемая селективность к определенному виду ионов. Электроды с жидкой и пленочной мембраной Жидкие мембраны это растворы в органических растворителях ионообменных веществ жидкие катиониты или аниониты или нейтральных хелатов отделенные от водных растворов нейтральными пористыми перегородками полимерными стеклянными или др. В настоящее время промышленность выпускает пленочные ионоселективные электроды на катионы N К NH4 Са2... | |||
Если основная роль липидов в составе мембран заключается в стабилизации бислоя, то белки отвечают за функциональную активность мембран. Одни из них обеспечивают транспорт определённых молекул и ионов, другие являются ферментами, третьи участвуют в связывании цитоскелета с внеклеточным матриксом или служат рецепторами для гормонов, медиаторов,
эйкозаноидов, липопротеинов, оксида азота (N0). На долю белков приходится от 30 до 70% массы мембран. Белки определяют особенности функционирования каждой мембраны.
Особенности строения
и локализации белков в мембранах
Мембранные белки, контактирующие с гидрофобной частью липидного бислоя, должны быть амфифильными. Те участки белка, которые взаимодействуют с углеводородными цепями жирных кислот, содержат преимущественно неполярные аминокислоты. Участки белка, находящиеся в области полярных «головок», обогащены гидрофильными аминокислотными остатками.
Локализация белков в мембранах. Трансмембранные белки, например: 1 - гликофорин А; 2 - рецептор адреналина. Поверхностные белки: 3 - белки, связанные с интегральными белками, например, фермент сукцинатдегидрогеназа; 4 - белки, присоединенные к полярным «головкам» липидного слоя, например, протеинкинаэа С; 5 - белки, -заякоренные» в мембране с помощью короткого гидрофобного концевого домена, например, цитохрои b 5 ;6 - «заякоренные» белки, ковалентно соединённые с пипидом мембраны (например, фермент щелочная фосфатаза).
Белки мембран различаются по своему положению в мембране. Они могут глубоко проникать в липидный бислой или даже пронизывать его - интегральные белки, либо разными способами прикрепляться к мембране - поверхностные белки.Поверхностные белки
Поверхностные белки часто прикрепляются к мембране, взаимодействуя с интегральными
белками или поверхностными участками липидного слоя.
Белки, образующие комплексы с интегральными белками мембраны
Ряд пищеварительных ферментов, участвующих в гидролизе крахмала и белков, прикрепляется к интегральным белкам мембран микроворсинок кишечника.
Примерами таких комплексов могут быть сахараза-изомальтаза и мальтаза-гликоамилаза.
Белки, связанные с полярными «головками» липидов мембран
Полярные или заряженные домены белковой молекулы могут взаимодействовать с полярными «головками» липидов, образуя ионные и водородные связи. Кроме того, множество растворимых в цитозоле белков при определённых условиях могут связываться с поверхностью мембраны на непродолжительное время. Иногда связывание белка - необходимое условие проявления ферментативной активности. К таким белкам, например, относят протеинкиназу С, факторы свёртывания крови.
Закрепление с помощью мембранного «якоря»
«Якорем» может быть неполярный домен белка, построенный из аминокислот с гидро-
фобными радикалами. Примером такого белка может служить цитохром b 5 мембраны ЭР. Этот белок участвует в окислительно-восстановительных реакциях, как переносчик электронов.
Роль мембранного «якоря» может выполнять также ковалентно связанный с белком остаток жирной кислоты (миристиновой - С 14 или пальмитиновой - С 16). Белки, связанные с жирными кислотами, локализованы в основном на внутренней поверхности плазматической мембраны. Миристиновая кислота присоединяется к N-концевому глицину с образованием амидной связи. Пальмитиновая кислота образует тиоэфирную связь с цистеином или слож-ноэфирную с остатками серина и треонина.
Небольшая группа белков может взаимодействовать с наружной поверхностью клетки с помощью ковалентно присоединённого к С-концу белка фосфатидилинозитолгликана. Этот «якорь» - часто единственное связующее звено между белком и мембраной, поэтому при действии фосфолипазы С этот белок отделяется от мембраны.
Трансмембранные (интегральные) белки
Некоторые из трансмембранных белков пронизывают мембрану один раз (гликофорин), другие имеют несколько участков (доменов), последовательно пересекающих бислой.
Трансмембранные домены, пронизывающие бислой, имеют конформацию α -спирали. Полярные остатки аминокислот обращены внутрь глобулы, а неполярные контактируют с мембранными липидами. Такие белки называют «вывернутыми» по сравнению с растворимыми в воде белками, в которых большинство гидрофобных остатков аминокислот спрятано внутрь, а гидрофильные располагаются на поверхности.
Радикалы заряженных аминокислот в составе этих доменов лишены заряда и протониро-ваны (-СООН) или депротонированы (-NH 2).
Гликозилированные белки
Поверхностные белки или домены интегральных белков, расположенные на наружной поверхности всех мембран, почти всегда гликози-лированы. Олигосахаридные Остатки могут быть присоединены через амидную группу аспараги-на или гидроксильные группы серина и треонина.
Олигосахаридные остатки защищают белок от протеолиза, участвуют в узнавании лигандов или адгезии.
Латеральная диффузия белков
Некоторые мембранные белки перемещаются вдоль бислоя (латеральная диффузия) или поворачиваются вокруг оси, перпендикулярно его поверхности.
Латеральная диффузия интегральных белков в мембране ограничена, это связано с их большими размерами, взаимодействием с другими мембранными белками, элементами цитоскелета или внеклеточного матрикса.
Белки мембран не совершают перемещений с одной стороны мембраны на другую («флип-флоп» перескоки), подобно фосфолипидам.
9.5.1. Одна из главных функций мембран - участие в переносе веществ. Этот процесс обеспечивается при помощи трёх основных механизмов: простой диффузией, облегчённой диффузией и активным транспортом (рисунок 9.10). Запомните важнейшие особенности этих механизмов и примеры транспортируемых веществ в каждом случае.
Рисунок 9.10. Механизмы транспорта молекул через мембрану
Простая диффузия - перенос веществ через мембрану без участия специальных механизмов. Транспорт происходит по градиенту концентрации без затраты энергии. Путём простой диффузии транспортируются малые биомолекулы - Н2 О, СО2 , О2 , мочевина, гидрофобные низкомолекулярные вещества. Скорость простой диффузии пропорциональна градиенту концентрации.
Облегчённая диффузия - перенос веществ через мембрану при помощи белковых каналов или специальных белков-переносчиков. Осуществляется по градиенту концентрации без затраты энергии. Транспортируются моносахариды, аминокислоты, нуклеотиды, глицерол, некоторые ионы. Характерна кинетика насыщения - при определённой (насыщающей) концентрации переносимого вещества в переносе принимают участие все молекулы переносчика и скорость транспорта достигает предельной величины.
Активный транспорт - также требует участия специальных белков-переносчиков, но перенос происходит против градиента концентрации и поэтому требует затраты энергии. При помощи этого механизма через клеточную мембрану транспортируются ионы Na+ , K+ , Ca2+ , Mg2+ , через митохондриальную - протоны. Для активного транспорта веществ характерна кинетика насыщения.
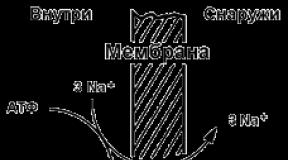
9.5.2. Примером транспортной системы, осуществляющей активный транспорт ионов, является Na+ ,K+ -аденозинтрифосфатаза (Na+ ,K+ -АТФаза или Na+ ,K+ -насос). Этот белок находится в толще плазматической мембраны и способен катализировать реакцию гидролиза АТФ. Энергия, выделяемая при гидролизе 1 молекулы АТФ, используется для переноса 3 ионов Na+ из клетки во внеклеточное пространство и 2 ионов К+ в обратном направлении (рисунок 9.11). В результате действия Na+ ,K+ -АТФазы создаётся разность концентраций между цитозолем клетки и внеклеточной жидкостью. Поскольку перенос ионов неэквивалентен, то возникает разность электрических потенциалов. Таким образом, возникает электрохимический потенциал, который складывается из энергии разности электрических потенциалов Δφ и энергии разности концентраций веществ ΔС по обе стороны мембраны.

Рисунок 9.11. Схема Na+ , K+ -насоса.
9.5.3. Перенос через мембраны частиц и высокомолекулярных соединений
Наряду с транспортом органических веществ и ионов, осуществляемым переносчиками, в клетке существует совершенно особый механизм, предназначенный для поглощения клеткой и выведения из неё высокомолекулярных соединений при помощи изменения формы биомембраны. Такой механизм называют везикулярным транспортом .

Рисунок 9.12. Типы везикулярного транспорта: 1 - эндоцитоз; 2 - экзоцитоз.
При переносе макромолекул происходит последовательное образование и слияние окружённых мембраной пузырьков (везикул). По направлению транспорта и характеру переносимых веществ различают следующие типы везикулярного транспорта:
Эндоцитоз (рисунок 9.12, 1) — перенос веществ в клетку. В зависимости от размера образующихся везикул различают:
а) пиноцитоз — поглощение жидкости и растворённых макромолекул (белков, полисахаридов, нуклеиновых кислот) с помощью небольших пузырьков (150 нм в диаметре);
б) фагоцитоз — поглощение крупных частиц, таких, как микроорганизмы или обломки клеток. В этом случае образуются крупные пузырьки, называемые фагосомами диаметром более 250 нм.
Пиноцитоз характерен для большинства эукариотических клеток, в то время как крупные частицы поглощаются специализированными клетками - лейкоцитами и макрофагами. На первой стадии эндоцитоза вещества или частицы адсорбируются на поверхности мембраны, этот процесс происходит без затраты энергии. На следующей стадии мембрана с адсорбированным веществом углубляется в цитоплазму; образовавшиеся локальные впячивания плазматической мембраны отшнуровываются от поверхности клетки, образуя пузырьки, которые затем мигрируют внутрь клетки. Этот процесс связан системой микрофиламентов и является энергозависимым. Поступившие в клетку пузырьки и фагосомы могут сливаться с лизосомами. Содержащиеся в лизосомах ферменты расщепляют вещества, содержащиеся в пузырьках и фагосомах до низкомолекулярных продуктов (аминокислот, моносахаридов, нуклеотидов), которые транспортируются в цитозоль, где они могут быть использованы клеткой.
Экзоцитоз (рисунок 9.12, 2) — перенос частиц и крупных соединений из клетки. Этот процесс, как и эндоцитоз, протекает с поглощением энергии. Основными разновидностями экзоцитоза являются:
а) секреция - выведение из клетки водорастворимых соединений, которые используются или воздействуют на другие клетки организма. Может осуществляться как неспециализированными клетками, так и клетками эндокринных желёз, слизистой желудочно-кишечного тракта, приспособленными для секреции производимых ими веществ (гормонов, нейромедиаторов, проферментов) в зависимости от определённых потребностей организма.
Секретируемые белки синтезируются на рибосомах, связанных с мембранами шероховатого эндоплазматического ретикулума. Затем эти белки транспортируются к аппарату Гольджи, где они модифицируются, концентрируются, сортируются, и затем упаковываются в пузырьки, которые отщепляются в цитозоль и в дальнейшем сливаются с плазматической мембраной, так что содержимое пузырьков оказывается вне клетки.
В отличие от макромолекул, секретируемые частицы малых размеров, например, протоны, транспортируются из клетки при помощи механизмов облегчённой диффузии и активного транспорта.
б) экскреция - удаление из клетки веществ, которые не могут быть использованы (например, удаление в ходе эритропоэза из ретикулоцитов сетчатой субстанции, представляющей собой агрегированные остатки органелл). Механизм экскреции, по-видимому, состоит в том, что вначале выделяемые частицы оказываются в цитоплазматическом пузырьке, который затем сливается с плазматической мембраной.
Биологическая химия Лелевич Владимир Валерьянович
Белки мембран.
Белки мембран.
Мембранные белки отвечают за функциональную активность мембран и на их долю приходится от 30 до 70%. Белки мембран отличаются по своему положению в мембране. Они могут глубоко проникать в липидный бислой или даже пронизывать его – интегральные белки, разными способами прикрепляться к мембране – поверхностные белки, либо, ковалентно контактировать с ней – заякоренные белки. Поверхностные белки почти всегда гликозилированы. Олигосахаридные остатки защищают белок от протеолиза, участвуют в узнавании лигандов и адгезии.
Белки, локализованные в мембране, выполняют структурную и специфические функции:
1. транспортную;
2. ферментативную;
3. рецепторную;
4. антигенную.
Из книги Заводи кого угодно, только НЕ КРОКОДИЛА! автора Орсаг МихайНу а белки? В шестидесятых годах я неоднократно пытался завести в доме и белок, но каждая такая попытка кончалась самым печальным образом. Через некоторое время белки слабели, задние конечности у них отнимались и несчастные животные в судорогах погибали. Поначалу я
Из книги Геном человека: Энциклопедия, написанная четырьмя буквами автора Из книги Геном человека [Энциклопедия, написанная четырьмя буквами] автора Тарантул Вячеслав ЗалмановичНе все гены кодируют белки Прежде всего, следует отметить, что кроме генов, кодирующих белки, в геноме имеются еще гены, на которых синтезируется РНК, которые не являются мРНК (то есть, не кодируют белок), но выполняют ряд самостоятельных важных функций в клетках. В
Из книги Живые часы автора Уорд Ритчи11. Белки в колесе Чтобы познакомиться с современными поисками решения проблемы живых часов, обратимся к исследованиям, проводившимся биологами в последние годы. В числе первых следует, пожалуй, назвать работу Патриции де Курси.В 1955 году де Курси получила диплом
Из книги Биология [Полный справочник для подготовки к ЕГЭ] автора Лернер Георгий Исаакович Из книги Кровь: река жизни [От древних легенд до научных открытий] автора Азимов АйзекГлава 11 Эти подвижные белки В начале предыдущей главы я упомянул, что органические компоненты пищи делятся на три группы. Я рассказал об одной из этих групп: углеводах. Далее логично было бы перейти к белкам, потому что их метаболизм в организме происходит параллельно с
Из книги Логика случая [О природе и происхождении биологической эволюции] автора Кунин Евгений ВикторовичГлава 12 Происхождение жизни. Возникновение трансляции, репликации, метаболизма и мембран: биологический, геохимический и космологический подходы Пер. А. НеизвестногоВ предыдущей главе мы обсудили возможные сценарии возникновения клеток и (будем надеяться) достигли
Из книги Гены и развитие организма автора Нейфах Александр Александрович2. Белки хроматина Мы уже знаем, что хроматин состоит из ДНК и гистонов в равном весовом количестве и негистоновых белков (НГБ), которых в неактивных районах хромосомы всего 0,2 веса ДНК, а в активных - более чем 1,2 (в среднем НГБ мепьше, чем ДНК). Мы знаем также, что гистоны
Из книги Мир животных автора Ситников Виталий Павлович Из книги Современное состояние биосферы и экологическая политика автора Колесник Ю. А.4.1. Образование мембран – основа начала жизни Рассматривая гигантское многообразие современных живых организмов, можно прийти к выводу, что существовало множество путей развития, берущих начало от реликтовых форм жизни. На самом деле исследования молекулярной эволюции
Из книги Секреты наследственности человека автора Афонькин Сергей ЮрьевичКлетки, белки и гены Жизнь есть способ существования белковых тел. Ф. Энгельс Наше тело является империей клеток, каждая из которых представляет собой миниатюрную фабрику для производства белков. Многие из этих важнейших макромолекул могут быть выделены из организма в
Из книги Антропология и концепции биологии автора Курчанов Николай АнатольевичБелки Белки имеют первостепенное значение в жизни организмов. Огромное разнообразие живых существ в значительной степени определяется различиями в составе имеющихся в их организме белков. Например, в организме человека их известно более 5 млн.Белки – это полимеры,
Из книги Биологическая химия автора Лелевич Владимир ВалерьяновичХимический состав мембран. Мембраны состоят из липидных и белковых молекул, относительное количество которых у разных мембран широко колеблется. Углеводы содержатся в форме гликопротеинов, гликолипидов и составляют 0,5%-10% веществ мембраны. Согласно жидкостно-мозаичной
Из книги автораЛипиды мембран. Мембранные липиды – амфифильные молекулы, т.е. в молекуле есть как гидрофильные группы (полярные головки), так и алифатические радикалы (гидрофобные хвосты), самопроизвольно формирующие бислой, в котором хвосты липидов обращены друг к другу. Толщина
Из книги автораБелки Пищевая ценность белка обеспечивается наличием незаменимых аминокислот, углеводородные скелеты которых не могут синтезироваться в организме человека, и они соответственно должны поступать с пищей. Они также являются основными источниками азота. Суточная
Из книги автораБелки мышечной ткани Выделяют три группы белков:1. миофибриллярные белки – 45 %;2. саркоплазматические белки – 35 %;3. белки стромы – 20 %.Миофибриллярные белки.К этой группе относятся:1. миозин;2. актин;3. актомиозин;а также так называемые регуляторные белки:4. тропомиозин;5.
1. Клеточная мембрана - это ультратонкая пленка на поверхности клетки или клеточной органеллы, состоящая из бимолекулярного слоя липидов с встроенными белками и полисахаридами.
Функции мембран:
барьерная - обеспечивает селективный, регулируемый пассивный и активный транспорт.
матричная - обеспечивает определенное взаимное расположение и ориентацию мембранных ферментов относительно субстратов с целью реализации их оптимального взаимодействия.
механическая - обеспечивает прочность и автономность клетки и внутриклеточных структур
2 .Моделью организации на сегодняшний день признана жидкостно-кристаллическая модель. Впервые ее предложили Сингер и Николсон в 1972 г. Согласно этой модели основу любой мембраны составляет двойной фосфолипидный слой. Молекулы фосфолипидов ориентированы так, что их гидрофильные головки выходят наружу и образуют внешнюю и внутреннюю поверхности мембраны, а их гидрофобные хвосты направлены к середине бимолекулярного слоя. Белки как бы плавают в липидном слое. Поверхностные белки располагаются на внешней и внутренней поверхностях мембраны, удерживаясь преимущественно за счет электростатическими силами. Интегральные белки могут пронизывать двойной слой насквозь. Такие белки являются главным компонентом, ответственным за избирательную проницаемость кл. мембраны.
Кроме фосфолипидов и белков в биологических мембранах содержатся и другие хим. соединения (холестерин, гликолипиды, гликопротеиды).
Функции мембранных белков. Некоторые мембранные белки участвуют в транспорте малых молекул через мембрану клетки, изменяя ее проницаемость. Мембранные транспортные белки можно подразделить на белки-каналы и белки-переносчики. Белки-каналы содержат внутренние заполненные водой поры, которые позволяют ионам (через ионные каналы) или молекулы воды перемещаться через мембрану. Многие ионные каналы специализируются на транспорте только одного иона; так калиевые и натриевые каналы часто различают эти сходные ионы и пропускают только один из них. Белки-переносчики связывают каждую переносимую молекулу или ион и могут осуществлять активный транспорт с использованием энергии АТФ. Также белки играют важную роль в сигнальных системах клеток, при иммунном ответе и в кл. цикле.
Функции мембранных углеводов. Углеводная часть гликолипидов и гликопротеинов плазматической мембраны всегда находится на наружной поверхности мембраны, контактируя с межклеточным веществом. Полисахариды наряду с белками выполняют роль антигенов при развитии кл. иммунитета. Участвуют в транспорте гликопротеинов, направляя их к месту назначения в клетке или на ее поверхности.
Главная функция мембранных липидов состоит в том, что они формируют бислойный матрикс, с которым взаимодействуют белки.
Латеральная диффузия - это хаотическое тепловое перемещение молекул липидов и белков в плоскости мембраны. При латеральной диффузии рядом расположенные молекулы липидов скачком меняются местами и вследствие таких последовательных перескоков из одного места в другое молекула перемещается вдоль поверхности мембраны. Среднее квадратичное перемещение молекул при диффузии за время t можно оценивать по формуле Эйнштейна: Sкв = 2кор(Dt), где D - коэф. лат. диффузии молекулы.
Латеральная диффузия интегральных белков в мембране ограничена, это связано с их большими размерами, взаимодействием с др. мембр. белками. Белки мембран не совершают перемещений с одной стороны мембраны на другую ("флип-флоп" перескоки), подобно фосфолипидам.
Перемещение молекул по поверхности мембраны клетки за время t определено экспериментально методом флуоресцентных меток - флуоресцирующих молекулярных групп. Оказалось что среднее квадратичное перемещение фосфолипидной молекулы за секунду по поверхности мембраны эритроцита соответствует расстоянию 5 мкм, что сравнимо с размерами клеток. Аналогичная величина для белковых молекул составила 0,2 мкм за сек.
Рассчитанные по формуле Эйнштейна коэффициенты лат. диффузии для липидов 6*10 (в -12 ст) кВ.м/с, для белков - порядка 10 (в - 14 ст) кВ.м/с.
Частота перескоков молекулы с одного места на другое вследствие лат. диффузии может быть найдена по формуле:
v = 2кор(3D/f),
где f - площадь, занимаемая одной молекулой на мембране.
Искусственная мембрана обычно представляет собой жесткую селективно-проницаемую перегородку, разделяющую массообменный аппарат на две рабочие зоны, в которых поддерживаются различные давления и составы разделяемой смеси. Мембраны могут быть выполнены в виде плоских листов, труб, капилляров и полых волокон. Мембраны выстраиваются в мембранные системы. Наиболее распространенные искусственные мембраны -- полимерные мембраны. При определённых условиях, преимущественно могут быть использованы керамические мембраны.Некоторые мембраны работают в широком диапазоне мембранных операций, таких, как микрофильтрация, ультрафильтрация, обратный осмос, первапорация, сепарация газа, диализ или хроматография. Способ применения зависит от типа функциональности включеной в мембрану, которые могут быть основаны на изоляции по размеру, химическом родстве или электростатике.
Липосомы. или фосфолипидные везикулы (пузырьки), получают обычно при набухании сухих фосфолипидов в воде или при впрыскивании раствора липидов в воду. При этом происходит самосборка бимолекулярной липидной мембраны. При этом все неполярные гидрофобные хвосты находятся внутри мембраны и ни один из них не соприкасается с полярными молекулами воды, однако чаще получаются сферические многоламеллярные липосомы, состоящие из нескольких бимолекулярных слоев - многослойные липосомы.
Отдельные бимолекулярные слои многослойной липосомы отделены водной средой. Толщина липидных слоев составляет, в зависимости от природы липидов 6,5 - 7,5 нм, а расстояние между ними - 1,5 - 2 нм. Диаметр многослойных липосом колеблется от 60 нм до 400 нм и более. Однослойные липосомы можно получить различными методами, например из суспензии многослойных липосом, если обрапотать их ультразвуком. Диаметр однослойных липосом составляет 25 - 30 нм. Липосомы представляют собой в некотором роде прообраз клетки. Они служат моделью для исследований различных свойств кл. мембран. Липосомы нашли непосредственное применение и в медицине (фоффолипидная микрокапсула для доставки лекарства в определенные органы и ткани). Липосомы не токсичны, полностью усваиваются организмом.