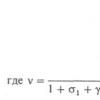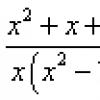От чего зависит скорость распространения нервного импульса. Нервные импульсы. Распространение нервного импульса по нервному волокну
В результате эволюции нервной системы человека и других животных возникли сложные информационные сети, процессы в которых основаны на химических реакциях. Важнейшим элементом нервной системы являются специализированные клетки нейроны . Нейроны состоят из компактного тела клетки, содержащего ядро и другие органеллы. От этого тела отходит несколько разветвленных отростков. Большинство таких отростков, называемых дендритами , служат точками контакта для приема сигналов от других нейронов. Один отросток, как правило самый длинный, называется аксоном и передает сигналы на другие нейроны. Конец аксона может многократно ветвиться, и каждая из этих более мелких ветвей способна соединиться со следующим нейроном.
Во внешнем слое аксона находится сложная структура, образованная множеством молекул, выступающих в роли каналов, по которым могут поступать ионы — как внутрь, так и наружу клетки. Один конец этих молекул, отклоняясь, присоединяется к атому-мишени. После этого энергия других частей клетки используется на то, чтобы вытолкнуть этот атом за пределы клетки, тогда как процесс, действующий в обратном направлении, вводит внутрь клетки другую молекулу. Наибольшее значение имеет молекулярный насос, который выводит из клетки ионы натрия и вводит в нее ионы калия (натрий-калиевый насос).
Когда клетка находится в покое и не проводит нервных импульсов, натрий-калиевый насос перемещает ионы калия внутрь клетки и выводит ионы натрия наружу (представьте себе клетку, содержащую пресную воду и окруженную соленой водой). Из-за такого дисбаланса разность потенциалов на мембране аксона достигает 70 милливольт (приблизительно 5% от напряжения обычной батарейки АА).
Однако при изменении состояния клетки и стимуляции аксона электрическим импульсом равновесие на мембране нарушается, и натрий-калиевый насос на короткое время начинает работать в обратном направлении. Положительно заряженные ионы натрия проникают внутрь аксона, а ионы калия откачиваются наружу. На мгновение внутренняя среда аксона приобретает положительный заряд. При этом каналы натрий-калиевого насоса деформируются, блокируя дальнейший приток натрия, а ионы калия продолжают выходить наружу, и исходная разность потенциалов восстанавливается. Тем временем ионы натрия распространяются внутри аксона, изменяя мембрану в нижней части аксона. При этом состояние расположенных ниже насосов меняется, способствуя дальнейшему распространению импульса. Резкое изменение напряжения, вызванное стремительными перемещения ионов натрия и калия, называют потенциалом действия . При прохождении потенциала действия через определенную точку аксона, насосы включаются и восстанавливают состояние покоя.
Потенциал действия распространяется довольно медленно — не более доли дюйма за секунду. Для того чтобы увеличить скорость передачи импульса (поскольку, в конце концов, не годится, чтобы сигнал, посланный мозгом, достигал руки лишь через минуту), аксоны окружены оболочкой из миелина, препятствующей притоку и оттоку калия и натрия. Миелиновая оболочка не непрерывна — через определенные интервалы в ней есть разрывы, и нервный импульс перескакивает из одного «окна» в другое, за счет этого скорость передачи импульса возрастает.
Когда импульс достигает конца основной части тела аксона, его необходимо передать либо следующему нижележащему нейрону, либо, если речь идет о нейронах головного мозга, по многочисленным ответвлениям многим другим нейронам. Для такой передачи используется абсолютно иной процесс, нежели для передачи импульса вдоль аксона. Каждый нейрон отделен от своего соседа небольшой щелью, называемой синапсом . Потенциал действия не может перескочить через эту щель, поэтому нужно найти какой-то другой способ для передачи импульса следующему нейрону. В конце каждого отростка имеются крошечные мешочки, называющие (пресинаптическими ) пузырьками , в каждом из которых находятся особые соединения — нейромедиаторы . При поступлении потенциала действия из этих пузырьков высвобождаются молекулы нейромедиаторов, пересекающие синапс и присоединяющиеся к специфичным молекулярным рецепторам на мембране нижележащих нейронов. При присоединении нейромедиатора равновесие на мембране нейрона нарушается. Сейчас мы рассмотрим, возникает ли при таком нарушении равновесия новый потенциал действия (нейрофизиологи продолжают искать ответ на этот важный вопрос до сих пор).
После того как нейромедиаторы передадут нервный импульс от одного нейрона на следующий, они могут просто диффундировать, или подвергнуться химическому расщеплению, или вернуться обратно в свои пузырьки (этот процесс нескладно называется обратным захватом ). В конце XX века было сделано поразительное научное открытие — оказывается, лекарства, влияющие на выброс и обратный захват нейромедиаторов, могут коренным образом изменять психическое состояние человека. Прозак (Prozac*) и сходные с ним антидепрессанты блокируют обратный захват нейромедиатора серотонина. Складывается впечатление, что болезнь Паркинсона взаимосвязана с дефицитом нейромедиатора допамина в головном мозге. Исследователи, изучающие пограничные состояния в психиатрии, пытаются понять, как эти соединения влияют на человеческий рассудок.
По-прежнему нет ответа на фундаментальный вопрос о том, что же заставляет нейрон инициировать потенциал действия — выражаясь профессиональным языком нейрофизиологов, неясен механизм «запуска» нейрона. В этом отношении особенно интересны нейроны головного мозга, которые могут принимать нейромедиаторы, посланные тысячей соседей. Об обработке и интеграции этих импульсов почти ничего не известно, хотя над этой проблемой работают многие исследовательские группы. Нам известно лишь, что в нейроне осуществляется процесс интеграции поступающих импульсов и выносится решение, следует или нет инициировать потенциал действия и передавать импульс дальше. Этот фундаментальный процесс управляет функционированием всего головного мозга. Неудивительно, что эта величайшая загадка природы остается, по крайней мере сегодня, загадкой и для науки!
Механизмы взаимодействия нервных клеток
Нервные клетки функционируют в тесном взаимодействии друг с другом.
Значение нервных импульсов. Все взаимодействия между нервными клетками осуществляются благодаря двум механизмам: 1) влияниям электрических полей нервных клеток (электротоническим влияниям) и 2) влияниям нервных импульсов.
Первые распространяются на очень небольшие территории мозга Электрический заряд нервной клетки создает вокруг нее электрическое поле, колебания которого вызывают изменения электрических полей лежащих рядом нейронов, что приводит к изменениям их возбудимости, лабильности и проводимости. Электрическое поле нейрона имеет сравнительно небольшую протяженность-около 100 мк, оно быстро затухает по мере удаления от клетки и может оказывать воздействие лишь на соседние нейроны.
Второй механизм обеспечивает не только ближайшие взаимодействия, но и передачу нервных влияний на большие расстояния. Именно с помощью нервных импульсов происходит объединение отдаленных и изолированных участков мозга в общую, синхронно работающую систему, что необходимо для протекания сложных форм деятельности организма. Нервный импульс, следовательно, является основным средством связи между нейронами. Высокая скорость распространения импульсов и локальное их воздействие на избранную точку мозга способствуют быстрой и точной передаче информации в нервной системе. В межнейронных взаимодействиях используется частотный код, т. е. изменения функционального состояния и характера ответных реакций одной нервной клетки кодируются изменением частоты импульсов (потенциалов действия), которые она посылает к другой нервной клетке. Общее количество импульсов, отправляемых нервной клеткой в единицу времени, или ее суммарная импульсная активность,-важный физиологический показатель деятельности нейрона.
Основные элементы химического синапса: синаптическая щель, везикулы (синаптические пузырьки), нейромедиаторы, рецепторы.
Си́напс (греч. σύναψις, от συνάπτειν - обнимать, обхватывать, пожимать руку) - место контакта между двумянейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Передача импульсов осуществляется химическим путём с помощью медиаторов или электрическим путём посредством прохождения ионов из одной клетки в другую.
Термин был введён в 1897 г. английским физиологом Чарльзом Шеррингтоном. Однако сам Шеррингтон утверждал, что получил идею этого термина в разговоре от физиолога Майкла Фостера .
Классификации синапсов

Основные элементы электрического синапса (эфапса): а - коннексон в закрытом состоянии; b - коннексон в открытом состоянии; с - коннексон, встроенный в мембрану; d - мономер коннексина, е -плазматическая мембрана; f - межклеточное пространство; g - промежуток в 2-4 нанометра в электрическом синапсе; h - гидрофильный канал коннексона.
По механизму передачи нервного импульса
химический - это место близкого прилегания двух нервных клеток, для передачи нервного импульса через которое клетка-источник выпускает в межклеточное пространство особое вещество, нейромедиатор, присутствие которого в синаптической щели возбуждает или затормаживает клетку-приёмник.
электрический (эфапс) - место более близкого прилегания пары клеток, где их мембраны соединяются с помощью особых белковых образований - коннексонов (каждый коннексон состоит из шести белковых субъединиц). Расстояние между мембранами клетки в электрическом синапсе - 3,5 нм (обычное межклеточное - 20 нм). Так как сопротивление внеклеточной жидкости мало(в данном случае), импульсы через синапс проходят не задерживаясь. Электрические синапсы обычно бывают возбуждающими.
смешанные синапсы - Пресинаптический потенциал действия создает ток, который деполяризуетпостсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны не плотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
Наиболее распространены химические синапсы. Для нервной системы млекопитающих электрические синапсы менее характерны, чем химические.
По местоположению и принадлежности структурам[править | править вики-текст]
нервно-мышечные
нейросекреторные (аксо-вазальные)
рецепторно-нейрональные
аксо-дендритические - с дендритами, в том числе
аксо-шипиковые - с дендритными шипиками, выростами на дендритах;
периферические
центральные
аксо-соматические - с телами нейронов;
аксо-аксональные - между аксонами;
дендро-дендритические - между дендритами;

Различные варианты расположения химических синапсов
По нейромедиатору
в том числе адренергические, содержащие адреналин или норадреналин;
аминергические, содержащие биогенные амины (например, серотонин, дофамин);
холинергические, содержащие ацетилхолин;
пуринергические, содержащие пурины;
пептидергические, содержащие пептиды.
При этом в синапсе не всегда вырабатывается только один медиатор. Обычно основной медиатор выбрасывается вместе с другим, играющим роль модулятора.
По знаку действия
возбуждающие
тормозные .
Если первые способствуют возникновению возбуждения в постсинаптической клетке (в них в результате поступления импульса происходит деполяризация мембраны, которая может вызвать потенциал действия при определённых условиях.), то вторые, напротив, прекращают или предотвращают его появление, препятствуют дальнейшему распространению импульса. Обычно тормозными являются глицинергические (медиатор - глицин) и ГАМК-ергические синапсы (медиатор - гамма-аминомасляная кислота).
Тормозные синапсы бывают двух видов: 1) синапс, в пресинаптических окончаниях которого выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала; 2) аксо-аксональный синапс, обеспечивающий пресинаптическое торможение. Синапс холинергический (s. cholinergica) - синапс, медиатором в котором является ацетилхолин.
В некоторых синапсах присутствует постсинаптическое уплотнение - электронно-плотная зона, состоящая из белков. По её наличию или отсутствию выделяют синапсы асимметричные и симметричные . Известно, что все глутаматергические синапсы асимметричны, а ГАМКергические - симметричны.
В случаях, когда с постсинаптической мембраной контактирует несколько синаптических расширений, образуются множественные синапсы .
К специальным формам синапсов относятся шипиковые аппараты , в которых с синаптическим расширением контактируют короткие одиночные или множественные выпячивания постсинаптической мембраны дендрита. Шипиковые аппараты значительно увеличивают количество синаптических контактов на нейроне и, следовательно, количество перерабатываемой информации. «Не-шипиковые» синапсы называются «сидячими». Например, сидячими являются все ГАМК-ергические синапсы.
Проведение нервных импульсов по нервным волокнам и через синапсы. Высоковольтный потенциал, возникающий при возбуждении рецептора в нервном волокне, в 5-10 раз больше порога раздражения рецептора. Проведение волны возбуждения по нервному волокну обеспечивается тем, что каждый последующий его участок раздражается высоковольтным потенциалом предыдущего участка. В мякотных нервных волокнах этот потенциал распространяется не непрерывно, а скачкообразно; он перескакивает через один или даже несколько перехватов Ранвье, в которых усиливается. Продолжительность проведения возбуждения между двумя соседними перехватами Ранвье равняется 5-10% длительности высоковольтного потенциала.
Проведение нервного импульса по нервному волокну происходит только при условии его анатомической непрерывности и нормального физиологического его состояния. Нарушение физиологических свойств нервного волокна сильным охлаждением или отравлением ядами и наркотиками прекращает проведение нервного импульса даже при анатомической его непрерывности.
Нервные импульсы проводятся изолированно по отдельным двигательным и чувствительным нервным волокнам, которые входят в состав смешанного нерва, что зависит от изолирующих свойств покрывающих их миелиновых оболочек. В безмякотных нервных волокнах биоток распространяется непрерывно вдоль волокна и благодаря соединительнотканой оболочке не переходит с одного волокна на другое. Нервный импульс может распространяться по нервному волокну в двух направлениях: центростремительном и центробежном. Следовательно, существуют три правила проведения нервного импульса в нервных волокнах: 1) анатомической непрерывности и физиологической целости, 2) изолированного проведения и 3) двустороннего проведения.
Через 2-3 дня после отделения нервных волокон от тела нейрона они начинают перерождаться, или дегенерировать, и проведение нервных импульсов прекращается. Нервные волокна и миелин разрушаются и сохраняется только соединительнотканая оболочка. Если соединить перерезанные концы нервных волокон, или нерва, то после дегенерации тех участков, которые отделены от нервных клеток, начинается восстановление, или регенерация, нервных волокон со стороны тел нейронов, из которых они прорастают в сохранившиеся соединительнотканые оболочки. Регенерация нервных волокон приводит к восстановлению проведения импульсов.
В отличие от нервных волокон через нейроны нервной системы нервные импульсы проводятся только в одном направлении - от рецептора к работающему органу. Это зависит от характера проведения нервного импульса через синапсы. В нервном волокне над пресинаптической мембраной есть множество мельчайших пузырьков ацетилхолина. При достижении биотоком пресинаптической мембраны часть этих пузырьков лопается, и ацетилхолин проходит через мельчайшие отверстия в пресинаптической мембране в синаптическую щель.
В постсинаптической мембране имеются участки, обладающие особым сродством к ацетилхолину, который вызывает временное появление пор в постсинаптической мембране, отчего она становится временно проницаемой для ионов. В результате в постсинаптической мембране возникает возбуждение и высоковольтный потенциал, который распространяется по следующему нейрону или по иннервируемому органу. Следовательно, передача возбуждения через синапсы происходит химическим путем посредством посредника, или медиатора, ацетилхолина, а проведение возбуждения по следующему нейрону снова осуществляется электрическим путем.
Действие ацетилхолина на проведение нервного импульса через синапс кратковременно; он быстро разрушается, гидролизуется ферментом холинэстеразой.
Так как химическая передача нервного импульса в синапсе происходит в течение доли мсек, то в каждом синапсе нервный импульс на это время задерживается.
В отличие от нервных волокон, в которых информация передается по принципу «все или ничего», т. е. дискретно, в синапсах информация передается по принципу «больше или меньше», т. е. градуально. Чем больше до некоторого предела образуется медиатора ацетилхолина, тем выше частота высоковольтных потенциалов в последующем нейроне. После этого предела возбуждение переходит в торможение. Таким образом, цифровая информация, передаваемая по нервным волокнам, переходит в синапсах в измерительную информацию. Измерительные электронные машины,
в которых имеются определенные соотношения между реально измеряемыми количествами и теми величинами, которые они представляют, называются аналоговыми, работающими по принципу «больше или меньше»; можно считать, что в синапсах происходит аналогичный процесс и совершается его переход в цифровой. Следовательно, нервная система функционирует по смешанному типу: в ней совершаются и цифровые и аналоговые процессы.
Нервная система контролирует, координирует и регулирует согласованную работу всех систем органов, поддержание постоянства состава его внутренней среды (благодаря этому организм человека функционирует как единое целое). При участии нервной системы осуществляется связь организма с внешней средой.
Нервная ткань
Нервная система образована нервной тканью , которая состоит из нервных клеток - нейронов и мелких клеток спутников (глиальных клеток ), которых примерно в 10 раз больше, чем нейронов.
Нейроны обеспечивают основные функции нервной системы: передачу, переработку и хранение информации. Нервные импульсы имеют электрическую природу и распространяются по отросткам нейронов.
Клетки спутники
выполняют питательную, опорную и защитную функции, способствуя росту и развитию нервных клеток.
Строение нейрона
Нейрон - основная структурная и функциональная единица нервной системы.
Структурно-функциональной единицей нервной системы является нервная клетка – нейрон . Его основными свойствами являются возбудимость и проводимость.
Нейрон состоит из тела и отростков .
Короткие, сильно ветвящиеся отростки - дендриты , по ним нервные импульсы поступают к телу нервной клетки. Дендритов может быть один или несколько.
Каждая нервная клетка имеет один длинный отросток - аксон , по которому импульсы направляются от тела клетки . Длина аксона может достигать нескольких десятков сантиметров. Объединяясь в пучки, аксоны образуют нервы .
Длинные отростки нервной клетки (аксоны) покрыты миелиновой оболочкой . Скопления таких отростков, покрытых миелином (жироподобным веществом белого цвета), в центральной нервной системе образуют белое вещество головного и спинного мозга.
Короткие отростки (дендриты) и тела нейронов не имеют миелиновой оболочки, поэтому они серого цвета. Их скопления образуют серое вещество мозга.
Нейроны соединяются друг с другом таким образом: аксон одного нейрона присоединяется к телу, дендритам или аксону другого нейрона. Место контакта одного нейрона с другим называется синапсом . На теле одного нейрона насчитывается 1200–1800 синапсов.
Синапс - пространство между соседними клетками, в котором осуществляется химическая передача нервного импульса от одного нейрона к другому.
Каждый синапс состоит из трёх отделов :
- мембраны, образованной нервным окончанием (пресинаптическая мембрана );
- мембраны тела клетки (постсинаптическая мембрана );
- синаптической щели между этими мембранами
В пресинаптической части синапса содержится биологически активное вещество (медиатор ), которое обеспечивает передачу нервного импульса с одного нейрона на другой. Под влиянием нервного импульса медиатор выходит в синаптическую щель, действует на постсинаптическую мембрану и вызывает возбуждение в теле клетки следующего нейрона. Так через синапс передается возбуждение от одного нейрона к другому.
Распространение возбуждения связано с таким свойством нервной ткани, как проводимость .
Типы нейронов
Нейроны различаются по форме
В зависимости от выполняемой функции выделяют следующие типы нейронов:
- Нейроны, передающие сигналы от органов чувств в ЦНС (спинной и головной мозг), называют чувствительными . Тела таких нейронов располагаются вне ЦНС, в нервных узлах (ганглиях). Нервный узел представляет собой скопление тел нервных клеток за пределами центральной нервной системы.
- Нейроны, передающие импульсы от спинного и головного мозга к мышцам и внутренним органам называют двигательными . Они обеспечивают передачу импульсов от ЦНС к рабочим органам.
- Связь между чувствительными и двигательными нейронами осуществляется с помощью вставочных нейронов через синаптические контакты в спинном и головном мозге. Вставочные нейроны лежат в пределах ЦНС (т.е. тела и отростки этих нейронов не выходят за пределы мозга).
Скопление нейронов в центральной нервной системе называется ядром (ядра головного, спинного мозга).
Спинной и головной мозг связаны со всеми органами нервами .
Нервы - покрытые оболочкой структуры, состоящие из пучков нервных волокон, образованных в основном аксонами нейронов и клетками нейроглии.
Нервы обеспечивают связь центральной нервной системы с органами, сосудами и кожным покровом.
НЕРВНЫЙ ИМПУЛЬС
- волна возбуждения, к-рая
распространяется по нервному волокну и служит для передачи информации от периферич.
рецепторных (чувствительных) окончаний к нервным центрам, внутри центр. нервной
системы и от неё к исполнительным аппаратам - мышцам и железам. Прохождение
Н. и. сопровождается переходными электрич. процессами, к-рые можно зарегистрировать
как внеклеточными, так и внутриклеточными электродами.
Генерацию, передачу и переработку Н. и. осуществляет
нервная система. Осн. структурным элементом нервной системы высших организмов
является нервная клетка, или нейрон, состоящий из тела клетки и многочисл. отростков
- дендритов (рис. 1). Один из отростков у нериферич. нейронов имеет большую
длину - это нервное волокно, или аксон, протяжённость к-рого ~ 1 м, а толщина
от 0,5 до 30 мкм. Различают два класса нервных волокон: мякотные (мие-линизированные)
и безмякотные. У мякотных волокон имеется миелиновая оболочка, образованная
спец. мембраной, к-рая подобно изоляции накручивается на аксон. Протяжённость
участков сплошной миелиновой оболочки составляет от 200 мкм до 1 мм, они прерываются
т. н. перехватами Ранвье шириной 1мкм. Миелиновая оболочка играет роль изоляции;
нервное волокно на этих участках пассивно, электрически активна только мембрана
в перехватах Ранвье. Безмякотные волокна не имеют изолир. участков; их структура
однородна по всей длине, а мембрана обладает электрич. активностью по всей поверхности.
Нервные волокна заканчиваются на телах или ден-дритах
др. нервных клеток, но отделены от них проме-
жутком шириной ~ 10 нм. Эта область контакта
двух клеток наз. синапсом. Входящая в синапс мембрана аксона наз. пресинаптической,
а соответствующая мембрана дендритов или мышцы - пост-синаптической (см. Клеточные
структуры)
.
В нормальных условиях по нервному волокну постоянно
бегут серии Н. и., возникающих на дендритах или теле клетки и распространяющихся
по аксону в направлении от тела клетки (аксон может проводить Н. и. в обоих
направлениях). Частота этих периодич. разрядов несёт информацию о силе вызвавшего
их раздражения; напр., при умеренной активности частота ~ 50-100 импульсов/с.
Существуют клетки, к-рые разряжаются с частотой ~ 1500 импульсов/с.
Скорость распространения Н. и. u
зависит от типа нервного волокна и его диаметра d
, u
~ d
1/2 . В тонких волокнах нервной системы человека u
~ 1 м/с, а в толстых волокнах u
~ 100-120 м/с.
Каждый Н. и. возникает в результате раздражения
тела нервной клетки или нервного волокна. Н. и. всегда имеет одни и те же характеристики
(форму и скорость) независимо от силы раздражения, т. е. при подпороговом раздражении
Н. и. не возникает совсем, а при надпороговым - имеет полную амплитуду.
После возбуждения наступает рефракторный период, в течение к-рого возбудимость нервного волокна снижена. Различают абс. рефракторный период, когда волокно нельзя возбудить никакими раздражителями, и относит. рефракторный период, когда возбуждение возможно, но его порог оказывается выше нормы. Абс. рефракторный период ограничивает сверху частоту передачи Н. и. Нервное волокно обладает свойством аккомодации, т. е. привыкает к постоянно действующему раздражению, что выражается в постепенном повышении порога возбудимости. Это приводит к снижению частоты Н. и. и даже к их полному исчезновению. Если сила раздражения нарастает медленно, то возбуждения может не произойти даже после достижения порога.
Рис.1. Схема строения нервной клетки .
Вдоль нервного волокна Н. и. распространяется
в виде волны электрич. потенциала. В синапсе происходит смена механизма распространения.
Когда Н. и. достигает пресинаптич. окончания, в синаптич. щель выделяется активное
хим. вещество - м е д и а т о р. Медиатор диффундирует через синаптич. щель
и меняет проницаемость постсинаптич. мембраны, в результате чего на ней возникает
потенциал, вновь генерирующий распространяющийся импульс. Так действует хим.
синапс. Встречается также электрич. синапс, когда след. нейрон возбуждается
электрически.
Возбуждение Н. и
. Физ. представления о
появлении электрич. потенциалов в клетках основаны на т. н. мембранной теории.
Клеточные мембраны разделяют растворы электролита разной концентрации и обладают
из-бират. проницаемостью для нек-рых ионов. Так, мембрана аксона представляет
собой тонкий слой липидов и белков толщиной ~ 7 нм. Её электрич. сопротивление
в состоянии покоя ~ 0,1 Ом. м 2 , а ёмкость ~ 10 мф/м 2 .
Внутри аксона высока концентрация ионов К + и мала концентрация ионов
Na + и Сl - , а в окружающей среде - наоборот.
В состоянии покоя мембрана аксона проницаема
для ионов К + . Из-за разницы концентраций C 0 K
во внеш. и С во внутр. растворах на мембране устанавливается калиевый мембранный
потенциал
![]()
где Т
- абс. темп-pa, е
-
электрона. На мембране аксона действительно наблюдается потенциал покоя ~ -60
мВ, соответствующий указанной ф-ле.
Ионы Na + и Сl - проникают
через мембрану. Для поддержания необходимого неравновесного распределения ионов
клетка использует систему активного транспорта, на работу к-рой расходуется
клеточная энергия. Поэтому состояние покоя нервного волокна не является термодинамически
равновесным. Оно стационарно благодаря действию ионных насосов, причём мембранный
потенциал в условиях разомкнутой цепи определяется из равенства нулю полного
электрич. тока.
Процесс нервного возбуждения развивается следующим
образом (см. также Биофизика
).Если пропустить через аксон слабый импульс
тока, приводящий к деполяризации мембраны,
то после снятия внеш. воздействия потенциал монотонно возвращается к исходному
уровню. В этих условиях аксон ведёт себя как пассивная электрич. цепь, состоящая
из конденсатора и пост. сопротивления.
Рис. 2. Развитие потенциала действия в нервном
во
локне: а
- подпороговое
(1
) и надпороговое (2
) раздражения;
б
-мембранный отклик; при над-пороговом раздражении проявляется полный
потен
циал действия; в
- ионный
ток, протекающий через
мембрану при
возбуждении; г
- аппроксимация
ионного
тока в простой аналитической модели
.

Если импульс тока превышает нек-рую пороговую
величину, потенциал продолжает изменяться и после выключения возмущения; потенциал
становится положительным и только потом возвращается к уровню покоя, причём
вначале даже несколько проскакивает его (область гиперполяризации, рис. 2).
Отклик мембраны при этом не зависит от возмущения; этот импульс наз. потенциалом
действия. Одновременно через мембрану течёт ионный ток, направленный сначала
внутрь, а потом наружу (рис. 2, в
).
Феноменологич. истолкование механизма возникновения
Н. и. было дано А. Л. Ходжкином (A. L. Hodg-kin) и А. Ф. Хаксли (A. F. Huxley)
в 1952. Полный ионный ток слагается из трёх составляющих: калиевого, натриевого
и тока утечки. Когда потенциал мембраны сдвигается на пороговую величину j*
(~ 20мВ), мембрана становится проницаемой для ионов Na + . Ионы Na +
устремляются внутрь волокна, сдвигая мембранный потенциал, пока он не достигнет
величины равновесного натриевого потенциала:
![]()
составляющего ~ 60 мВ. Поэтому полная амплитуда
потенциала действия достигает ~ 120 мВ. К моменту достижения макс. потенциала
в мембране начинает развиваться калиевая (и одновременно уменьшаться натриевая)
проводимость. В результате натриевый ток сменяется
на калиевый, направленный наружу. Этот ток соответствует уменьшению потенциала
действия.
Установлены эмпирич. ур-ния для описания натриевого
и калиевого токов. Поведение мембранного потенциала при пространственно однородном
возбуждении волокна определяется ур-нием:
где С
- ёмкость мембраны, I
- ионный
ток, слагающийся из калиевого, натриевого и тока утечки. Эти токи определяются
пост. эдс j K ,
j Na
и j l
и проводимостями g
K , g
Na и g l:
Величину g l
считают постоянной,
проводимости g
Na и g
K описывают с
помощью параметров m
, h
и п:
g
Na , g
K -
постоянные; параметры т, h
и п
удовлетворяют линейным ур-ниям

Зависимость коэф. a
и b от мембранного
потенциала j
(рис. 3) выбирают из условия наилучшего совпадения

Рис. 3. Зависимость коэффициентов
a
и
b
от мембран
ного потенциала
.
расчётных и измеряемых кривых I
(t
).
Этими же соображениями вызван выбор параметров. Зависимость стационарных значений
т, h
и п
от мембранного потенциала приведена на рис. 4. Существуют
модели с большим числом параметров. Т. о., мембрана нервного волокна представляет
собой нелинейный ионный проводник, свойства к-рого существенно зависят от электрич.
поля. Механизм генерации возбуждения изучен плохо. Ур-ния Ходжкина -Хаксли дают
лишь удачное эмпирич. описание явления, за к-рым нет конкретной физ. модели.
Поэтому важной задачей является изучение механизмов протекания электрич. тока
через мембраны, в частности через управляемые элект- рич. полем ионные каналы.

Рис. 4. Зависимость стационарных значений
т, h
и п
от мембранного
потенциала
.
Распространение Н. и
. Н. и. может распространяться
вдоль волокна без затухания и с пост. скоростью. Это связано с тем, что необходимая
для передачи сигнала энергия не поступает из единого центра, а черпается на
месте, в каждой точке волокна. В соответствии с двумя типами волокон существуют
два способа передачи Н. и.: непрерывный и сальтаторный (скачкообразный), когда
импульс движется от одного перехвата Ранвье к другому,
перепрыгивая через области миелиновой изоляции.
В случае немиелинизир. волокна распределение
мембранного потенциала j(x,
t
)определяется ур-нием:
где С
- ёмкость мембраны, приходящаяся
на единицу длины волокна, R
- сумма продольных (внутриклеточного и внеклеточного)
сопротивлений на единицу длины волокна, I
- ионный ток, протекающий через
мембрану волокна единичной длины. Электрич. ток I
является функционалом
от потенциала j,
к-рый зависит от времени t
и координаты х
. Эта зависимость определяется
ур-ниями (2) -(4).
Вид функционала I
специфичен для биологически
возбудимой среды. Однако ур-ние (5), если отвлечься от вида I
, имеет
более общий характер и описывает многие физ. явления, напр. процесс горения.
Поэтому передачу Н. и. уподобляют горению порохового шнура. Если в бегущем пламени
процесс поджигания осуществляется за счёт , то в Н. и. возбуждение
происходит при помощи т. н. локальных токов (рис. 5).

Рис. 5. Локальные токи, обеспечивающие распростра
нение
нервного импульса
.
Ур-ния Ходжкина - Хаксли для распространения
Н. и. решались численно. Полученные решения вместе с накопленными эксперим.
данными показали, что распространение Н. и. не зависит от деталей процесса возбуждения.
Качеств. картину распространения Н. и. можно получить при помощи простых моделей,
отражающих лишь общие свойства возбуждения. Такой подход позволил рассчитывать
скорость и форму Н. и. в однородном волокне, их изменение при наличии неоднород-ностей
и даже сложные режимы распространения возбуждения в активных средах, напр. в
сердечной мышце. Существует неск. матем. моделей подобного рода. Простейшая
из них такова. Ионный ток, протекающий через мембрану при прохождении Н. и.,
является знакопеременным: вначале он течёт внутрь волокна, а потом наружу. Поэтому
его можно аппроксимировать кусочно-постоянной ф-цией (рис. 2, г
). Возбуждение
происходит, когда мембранный потенциал сдвигается на пороговую величину j*.
В этот момент возникает ток, направленный внутрь волокна и равный по модулю
j"
. Спустя время t"
ток меняется на противоположный, равный j
"". Эта фаза продолжается
в течение времени ~ t"".
решение ур-ния (5) можно найти как ф-цию переменной t = х/
u, где u
- скорость распространения Н. и. (рис. 2, б)
.
В реальных волокнах время t"
достаточно велико, поэтому только оно определяет скорость u, для к-рой справедлива ф-ла: ![]() . Учитывая, что j
" ~ ~d, R ~ d
2 и С
~
d
, где d
- диаметр волокна, находим в согласии с экспериментом,
что u ~ d
1/2 . С помощью кусочно-постоянной аппроксимации находят форму потенциала
действия.
. Учитывая, что j
" ~ ~d, R ~ d
2 и С
~
d
, где d
- диаметр волокна, находим в согласии с экспериментом,
что u ~ d
1/2 . С помощью кусочно-постоянной аппроксимации находят форму потенциала
действия.
Ур-ние (5) для распространяющегося Н. и. в действительности
допускает два решения. Второе решение оказывается неустойчивым; оно даёт Н.
и. со значительно меньшей скоростью и амплитудой потенциала. Наличие второго,
неустойчивого, решения имеет аналогию в теории горения. При распространении
пламени с боковым теплоотводом также возможно возникновение неустойчивого режима.
Простую аналитич. модель Н. и. можно усовершенствовать, учитывая дополнит. детали.
При изменении сечения и при ветвлении нервных
волокон прохождение Н. и. может быть затруднено или даже полностью блокировано.
В расширяющемся волокне (рис. 6) скорость импульса по мере приближения к расширению
убывает, а после расширения начинает расти, пока не выйдет на новое стационарное
значение. Замедление Н. и. тем сильнее, чем больше разница в сечениях. При достаточно
большом расширении Н. и. останавливается. Существует критич. расширение волокна,
к-рое задерживает Н. и.
При обратном движении Н. и. (из широкого волокна
в узкое) блокирования не происходит, но изменение скорости носит противоположный
характер. При подходе к сужению скорость Н. и. увеличивается, а затем начинает
спадать до нового стационарного значения. На графике скорости (рис., 6а
)
получается своего рода петля .
Рие. 6. Прохождение нервных импульсов по расширя
ющемуся
волокну: а
- изменение скорости импульса в
зависимости
от его направления; б
-схематическое
изображение
расширяющегося волокна
.

Другой тип неоднородности - ветвление волокон.
В узле ветвления возможны разл. варианты прохождения и блокирования импульсов.
При несинхронном подходе Н. и. условие блокирования
зависит от временного сдвига. Если временной сдвиг между импульсами мал, то
они помогают друг другу проникнуть в широкое третье волокно. Если сдвиг достаточно
велик, то Н. и. мешают друг другу. Связано это с тем, что Н. и., подошедший
первым, но не сумевший возбудить третье волокно, частично переводит узел в рефракторное
состояние. Кроме того, возникает эффект синхронизации: по мере приближения Н.
и. к узлу их запаздывание друг относительно друга уменьшается.
Взаимодействие Н. и
. Нервные волокна в
организме объединены в пучки или нервные стволы, образующие подобие многожильного
кабеля. Все волокна в пучке представляют собой самостоят. линии связи, но имеют
один общий "провод" - межклеточную жидкость. Когда по любому из
волокон бежит Н. и., он создаёт в межклеточной жидкости электрич. поле, к-рое
влияет на мембранный потенциал соседних волокон. Обычно такое влияние пренебрежимо
мало и линии связи работают без взаимных помех, но оно проявляется в пато-логич.
и искусств. условиях. Обрабатывая нервные стволы спец. хим. веществами, удаётся
наблюдать не только взаимные помехи, но и передачу возбуждения в соседние волокна.
Известны эксперименты по взаимодействию двух
нервных волокон, помещённых в ограниченный объём внеш. раствора. Если по одному
из волокон бежит Н. и., то одновременно изменяется возбудимость второго волокна.
Изменение проходит три стадии. Вначале возбудимость второго волокна падает (повышается
порог возбуждения). Это уменьшение возбудимости опережает потенциал действия,
бегущий по первому волокну, и длится примерно до тех пор, пока потенциал в первом
волокне не достигнет максимума. Затем возбудимость растёт, эта стадия совпадает
по времени с процессом уменьшения потенциала в первом волокне. Возбудимость
ещё раз уменьшается, когда в первом волокне происходит небольшая гиперполяризация
мембраны.
При одноврем. прохождении Н. и. по двум волокнам
иногда удавалось достигнуть их синхронизации. Несмотря на то что собств. скорости
Н. и. в разных волокнах различны, при их
одноврем. возбуждении мог возникнуть коллективный Н. и. Если собств. скорости
были одинаковы, то коллективный импульс имел меньшую скорость. При заметном
отличии собств. скоростей коллективная скорость имела промежуточное значение.
Синхронизоваться могли лишь Н. и., скорости к-рых отличались не слишком сильно.
Матем. описание этого явления даётся системой
ур-ний для мембранных потенциалов двух параллельных волокон j 1
и j 2:

где R
1 и R
2
- продольные сопротивления первого и второго волокон, R
3
- продольное сопротивление внешней среды, g
= R
1 R
2 + R
1 R
3
+ R
2 R
3 . Ионные токи I
1
и I
2 можно описать той или иной моделью нервного возбуждения.
При использовании простой аналитич. модели решение
приводит к след. картине. Когда возбуждается одно волокно, в соседнем наводится
знакопеременный мембранный потенциал: вначале волокно гиперполяри-зуется, затем
деполяризуется и, наконец, ещё раз ги-перполяризуется. Эти три фазы соответствуют
понижению, повышению и новому понижению возбудимости во-локна. При нормальных
значениях параметров сдвиг мембранного потенциала во второй фазе в сторону деполяризации
не достигает порога, поэтому передачи возбуждения в соседнее волокно не происходит.
При одноврем. возбуждении двух волокон система (6) допускает совместное автомодельное
решение, к-рое соответствует двум Н. и., движущимся с одинаковой скоростью на
пост. расстоянии друг от друга. Если впереди находится медленный Н. и., то он
притормаживает быстрый импульс, не выпуская его вперёд; оба движутся с относительно
малой скоростью. Если же впереди находится быстрый II. и., то он подтягивает
за собой медленный импульс. Коллективная скорость оказывается близкой к собств.
скорости быстрого импульса. В сложных нейронных структурах возможно появление
автоволи
.
Возбудимые среды
. Нервные клетки в организме
объединены в нейронные сети, к-рые в зависимости от частоты ветвления волокон
разделяют на редкие и густые. В редкой сети отд. элементы возбуждаются независимо
друг от друга и взаимодействуют только в узлах ветвления, как описано выше.
В густой сети возбуждение охватывает сразу много
элементов, так что их детальная структура и способ соединения между собой оказываются
несущественными. Сеть ведёт себя как непрерывная возбудимая среда, параметры
к-рой определяют возникновение и распространение возбуждения.
Возбудимая среда может быть трёхмерной, хотя
чаще её рассматривают как двумерную поверхность. Возбуждение, возникшее в к--л.
точке поверхности, распространяется во все стороны в виде кольцевой волны. Волна
возбуждения может огибать препятствия, но не может от них отражаться, не отражается
она и от границы среды. При столкновении волн между собой происходит их взаимное
уничтожение; пройти друг сквозь друга эти волны не могут из-за наличия позади
фронта возбуждения рефракторной области.
Примером возбудимой среды является сердечный
нервно-мышечный синцитий - объединение нервных и мышечных волокон в единую проводящую
систему, способную передавать возбуждение в любом направлении. Нервно-мышечные
синцитии сокращаются синхронно, подчиняясь волне возбуждения, к-рую посылает
единый управляющий центр - водитель ритма. Единый ритм иногда нарушается, возникают
аритмии. Один из таких режимов наз. трепетанием предсердий: это автономные сокращения,
вызванные циркуляцией возбуждения вокруг препятствия, напр.
верхней или нижней вены. Для возникновения подобного режима периметр препятствия
должен превышать длину волны возбуждения, равную в предсердии человека ~ 5 см.
При трепетании происходит пе-риодич. сокращение предсердий с частотой 3-5 Гц.
Более сложный режим возбуждения представляет собой фибрилляция желудочков сердца,
когда отд. элементы сердечной мышцы начинают сокращаться без внеш. команды и
без связи с соседними элементами с частотой ~ 10 Гц. Фибрилляция приводит к
прекращению циркуляции крови.
Возникновение и поддержание спонтанной активности
возбудимой среды неразрывно связаны с возникновением источников волн. Простейший
источник волн (группа спонтанно возбуждающихся клеток) может обеспечить периодич.
пульсацию активности, так устроен водитель ритма сердца.
Источники возбуждения могут возникать и за счёт
сложной пространств. организации режима возбуждения, напр. ревербератор типа
вращающейся спиральной волны, появляющийся в простейшей возбудимой среде. Другой
вид ревербератора возникает в среде, состоящей из элементов двух типов с разными
порогами возбуждения; ревербератор периодически возбуждает то одни, то другие
элементы, меняя при этом направление своего движения и порождая .
Третий вид источника - ведущий центр (источник
эха), к-рый появляется в среде, неоднородной по реф-ракторности или порогу возбуждения.
В этом случае на неоднородности возникает отражённая волна (эхо). Наличие подобных
источников волн приводит к появлению сложных режимов возбуждения, исследуемых
в теории автоволн.
Лит.: Ходжкин А., Нервный импульс , пер. с англ., М., 1965; Катц Б., Нерв, мышца и синапс, пер. с англ., М., 1968; Ходоров Б. И., Проблема возбудимости, Л., 1969; Тасаки И., Нервное возбуждение, пер. с англ., М., 1971; Маркин В. С., Пастушенко В. Ф., Чизмад-жев Ю. А., Теория возбудимых сред, М., 1981. В. С. Маркин .