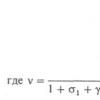Когнитивные функции локализация в коре головного мозга. Динамическая локализация функций в коре полушарий большого мозга. Топическая диагностика корковых поражений
Кора больших полушарий - материальная основа психической деятельности человека. Кора - серое вещество толщиной от 1,5 до 5 мм, содержит 14 млрд нервных клеток и имеет шестислойное строение. Кора - огромный ядерный центр, ядро, распластанное по поверхности полушарий.
Более 130 лет идет спор - есть ли в коре центры или нет и в каком объеме они оказывают влияние на «курируемые» функции: 1. Отвечают ли эти центры буквально за все (центр туризма, любви к живописи, к театру и пр.), или их влияние менее детализировано. 2. Кора - это один сплошной экранный центр, отвечающий за все функции.
Очевидно, истина, как всегда, где-то посередине.
Основоположником детального изучения клеточного состава коры был русский ученый, киевлянин Владимир Алексеевич Бец. В 1874 г. он опубликовал результат исследований с помощью собственного метода серийных срезов и окраски кармином. Бец выявил различное строение коры в различных ее участках и разработал карту цитоархитектоники коры. В последующем были созданы и другие карты: Бродмана с 52 цитоархитектоническими полями, Фогта со 150 миелоархитектоническими полями и др. Исследования в настоящнее время продолжаются в Институте мозга в Москве и в других странах.
Представления о локализации функций в коре головного мозга имеют большое практическое значение для решения задач топики поражений в полушариях мозга. Повседневный клинический опыт показывает, что существуют определенные закономерности зависимости расстройств функций от расположения патологического очага. Исходя из этого, клиницист и решает задачи топической диагностики. Однако так дело обстоит с простыми функциями: движением и чувствительностью. Функции более сложные, филогенетически молодые, не могут быть узколокализованными; в осуществлении сложных функций участвуют весьма обширные области коры, даже вся кора.
Работы В.А. Беца были внимательно изучены И.П. Павловым. С учетом этих данных, Иваном Петровичем Павловым были созданы основы нового и прогрессивного учения о локализации функций в головном мозге. Павлов рассматривал кору полушарий большого мозга как совокупность корковых концов анализаторов. Павлов создал учение об анализаторах. По Павлову, анализатор - нервный механизм, анализирующий явления внешнего и внутреннего мира путем разложения сложного комплекса раздражений на отдельные элементы. Он начинается воспринимающим аппаратом и кончается в мозгу, то есть анализатор включает рецепторный аппарат, проводник нервных импульсов и корковый центр.
Павлов доказал, что корковый конец анализатора - это не строго очерченная зона. В нем есть ядро и рассеянные элементы. Ядро - место концентрации нервных клеток, где происходит высший анализ, синтез и интеграция. На его периферии, в рассеянных элементах, совершаются простые анализ и синтез. Площади рассеянных элементов соседних анализаторов перекрывают друг друга (рис.).
По Павлову - работа второй сигнальной системы неразрывно связана с функциями всех анализаторов, поэтому невозможно представить локализацию сложных функций второй сигнальной системы в ограниченных корковых полях. Павловым заложены основы учения о динамической локализации функций в коре. Представления о динамической локализации функций в коре предполагают возможность использования одних и тех же корковых структур в разнообразных сочетаниях для обслуживания различных сложных корковых функций. Так, ассоциативные пути объединяют анализаторы, способствуя высшей синтетической деятельности коры мозга. Сегодня ученые знают, что раздражение трансформируется в возбуждение, передающееся в корковый конец анализатора. Не ясно другое - где и как возбуждение трансформируется в ощущение? Какие структуры отвечают за это? Так, при раздражении зрительного поля в области шпорной борозды возникают «простые» галлюцинации в виде световых или цветовых пятен, искр, теней. Раздражение наружной поверхности затылочной доли дает «сложные» галлюцинации в виде фигур, движущихся предметов.
В двигательной зоне коры обнаружены клетки, дающие разряд импульсов на зрительные, слуховые, кожные раздражения, а в зрительной зоне коры выявлены нейроны, отвечающие электрическими разрядами на осязательные, звуковые, вестибулярные и обонятельные раздражители. Кроме того, были найдены нейроны, которые отвечают не только на «свой» раздражитель, как теперь говорят, раздражитель своей модальности, своего качества, но и на один-два чужих. Их назвали полисенсорными нейронами.
Данный раздел анатомии НС разделен на следующие подкатегории
Лимбическая система - это функциональное объединение структур мозга, которое обеспечивает сложные формы поведения.
К лимбической системе относятся структуры древней коры, старой коры, мезокортекс и некоторые подкорковые образования. Особенностью лимбической системы является то, что связи между ее структурами образуют множество замкнутых кругов, и это создает условия для длительного циркулирования возбуждения в системе. Описаны основные круги, обладающие функциональной спецификой. Это большой круг Пейпса, который включает: гиппокамп - свод - мамиллярные тела - мамиллярно-таламический пучок Вик-д, Азира - передние ядра таламуса - кора поясной извилины - парагиппокампова извилина - гиппокамп.
Очень важной полифункциональной структурой в большом круге является гиппокамп. Его повреждение у человека нарушает память на события, которые предшествовали повреждению, нарушается запоминание, обработка новой информации, различение пространственных сигналов, снижается эмоциональность, инициативность, замедляется скорость протекания основных нервных процессов.
Малый круг Наута образуют: миндалина - конечная полоска - гипоталамус - перегородка - миндалина.
Важной структурой малого круга является миндалина. Ее функции связаны с обеспечением оборонительного поведения, вегетативными, двигательными, эмоциональными реакциями, мотивацией условнорефлекторного поведения. Многочисленные вегетативные эффекты миндалины обусловлены связью с гипоталамусом.
В целом лимбическая система обеспечивает:
- 1. Организацию вегетативно-соматических компонентов эмоций.
- 2. Организацию кратковременной и долговременной памяти.
- 3. Участвует в формировании ориентировочно-исследовательской деятельности (синдром Клювера - Бьюси).
- 4. Организует простейшую мотивационно - информационную коммуникацию (речь).
- 5. Участвует в механизмах сна.
- 6. Здесь находится центр обонятельной сенсорной системы.
По Мак Лину (1970) с функциональной точки зрения лимбика делится на: 1) нижний отдел - миндалина и гиппокамп, которые являются центрами эмоций и поведения для выживания и самосохранения; 2) верхний отдел - поясная извилина и височная кора, они представляют центры общительности и сексуальности; 3) средний отдел - гипоталамус и поясная извилина - центры биосоциальных инстинктов.
Полушария мозга состоят из белого вещества, которое снаружи покрыто серым веществом или корой. Кора самый молодой и сложный отдел мозга, где идет обработка сенсорной информации, формируются двигательные команды, интегрируются сложные формы поведения. Кроме нейронов, здесь имеется огромное количество глиальных клеток, которые выполняют ионорегулирующую и трофическую функцию.
Кора головного мозга имеет морфофункциональные особенности: 1) многослойность расположения нейронов; 2) модульный принцип организации; 3) соматотопическая локализация рецепторных систем; 4) экранность - распределение внешней рецепции на плоскости нейронального поля коркового конца анализатора; 5) зависимость уровня активности от влияния подкорковых структур и ретикулярной формации; 6) наличие представительства всех функций нижележащих структур ЦНС; 7) цитоархитектоническое распределение на поля; 8) наличие в специфических проекционных сенсорных и моторной системах коры вторичных и третичных полей с превалированием ассоциативных функций; 9) наличие специализированных ассоциативных областей коры; 10) динамическая локализация функций, которая выражается в возможности компенсации функций утраченных структур коры; 11) перекрытие в коре зон соседних периферических рецептивных полей; 12) возможность длительного сохранения следов раздражения; 13) реципрокная функциональная взаимосвязь возбудительных и тормозных состояний коры; 14) способность к иррадиации состояния; 15) наличие специфической электрической активности.
Кора состоит из 6 слоев:
- 1. Наружный молекулярный слой представлен сплетением нервных волокон, которые лежат параллельно поверхности корковых извилин и являются в основном дендритами пирамидных клеток. Сюда приходят афферентные таламокортикальные волокна от неспецифических ядер таламуса, они регулируют уровень возбудимости корковых нейронов.
- 2. Наружный зернистый слой образован мелкими звездчатыми клетками, которые определяют длительность циркулирования возбуждения в коре и имеют отношение к памяти.
- 3. Наружный пирамидный слой образуют средние по величине пирамидные клетки.
Функционально 2-й и 3-й слои осуществляют кортико-кортикальные ассоциативные связи.
- 4. К внутреннему зернистому слою приходят афферентные таламокортикальные волокна от специфических (проекционных) ядер таламуса.
- 5. Внутренний пирамидный слой образуют гигантские пирамидные клетки Беца. Аксоны этих клеток образуют кортикоспинальные и кортикобульбарные тракты, которые участвуют в координации целенаправленных движений и позы.
- 6. Полиморфный или слой веретеновидных клеток. Здесь формируются кортикоталамические пути.
Для всех анализаторов характерен соматотопический принцип организации проекции на кору периферических рецепторных систем. Например, в сенсорной коре II центральной извилины имеются участки представительства каждой точки кожной поверхности, в двигательной коре каждая мышца имеет свою топику, свое место, в слуховой коре имеется топическая локализация определенных тонов.
Особенностью корковых полей является экранный принцип функционирования, который заключается в том, что рецептор проецирует свой сигнал не на один нейрон коры, а на их поле, которое образуется коллатералями и связями нейронов. Сигнал при этом фокусируется не точка в точку, а на множестве нейронов, что и обеспечивает его полный анализ и возможность, при необходимости, передачи в другие структуры.
В вертикальном направлении входные и выходные волокна вместе со звездчатыми клетками образуют «колонки», которые являются функциональными единицами коры. И при погружении микроэлектрода перпендикулярно в кору, он на всем пути встречает нейроны, реагирующие на один вид раздражения, в то время если микроэлектрод идет горизонтально по коре, то он встречает нейроны, реагирующие на разные виды стимулов.
Наличие структурно различных полей предполагает и разное их функциональное назначение.
Важнейшая двигательная область коры находится в прецентральной извилине. В 30 г.г. прошлого столетия Пенфилд установил наличие правильной пространственной проекции соматических мышц различных отделов тела на двигательную область коры. Наиболее обширными и с самым низким порогом являются зоны, управляющие движениями кистей рук и мимической мускулатурой лица. На медиальной поверхности рядом с первичной была обнаружена вторичная двигательная область. Но эти области помимо моторного выхода из коры имеют самостоятельные сенсорные входы от кожных и мышечных рецепторов, поэтому их назвали первичной и вторичной мотосенсорной корой.
В постцентральной извилине находится первая соматосенсорная область, куда приходят афферентные сигналы от специфических ядер таламуса. Они несут информацию от рецепторов кожи и двигательного аппарата. И здесь отмечается соматотопическая организация.
Вторая соматосенсорная область находится в сильвиевой борозде, а т.к. первая и вторая соматосенсорная зона кроме афферентных входов имеет и моторные выходы их правильнее называть первичной и вторичной сенсомоторными зонами.
В затылочной области локализована первичная зрительная область.
В височной доле - слуховая область.
В каждой доле коры полушарий рядом с проекционными зонами расположены поля, которые не связаны с выполнением специфической функции --это ассоциативная кора, нейроны которой отвечают на раздражения различных модальностей и участвуют в интеграции сенсорной информации, а также обеспечивают связь между чувствительными и двигательными зонами коры. Это является физиологической основой высших психических функций.
Лобные доли имеют обширные двусторонние связи с лимбической системой мозга и участвуют в управлении врожденными поведенческими актами при помощи накопленного опыта, обеспечивают согласование внешних и внутренних мотиваций поведения, разработку стратегии поведения и программы действий, мыслительные особенности личности.
В деятельности полушарий нет полной симметрии. Так, у 9 из 10 человек, левое полушарие доминирует для двигательных актов (праворукие) и речи. У большинства «левшей» центр речи также находится слева. Т.е. абсолютного доминирования нет. Асимметрия полушарий особенно заметна при отделении одного полушария от другого (комиссуротомия). В левом полушарии находится центр письменной речи, стереогнозис. В левом полушарии лучше узнаются стимулы словесные, легко различимые, знакомые. Левым полушарием лучше выполняются задачи на временные отношения, установление сходства, идентичность стимулов по названиям. Левое полушарие осуществляет аналитическое и последовательное восприятие, обобщенное узнавание.
В правом полушарии осуществляется стереогнозис для левой руки, понимание элементарной речи, невербальное мышление (т.е. мышление образами), лучше узнаются стимулы несловесные, трудно различимые, незнакомые. Лучше выполняются задачи на пространственные отношения, установление различий, идентичность стимулов по физическим свойствам. В правом полушарии имеет место целостное, одновременное восприятие, конкретное узнавание.
Правое полушарие у 9 из 10 человек слегка приторможено, доминирует альфа-ритм, оно в свою очередь несколько притормаживает левое полушарие и не дает ему перевозбудиться. При выключении правого полушария человек много и непрерывно говорит (логорея), много обещает, но не выполняет обещания (болтун).
С усыплением левого полушария, наоборот, человек молчит, печален.
Правое полушарие отвечает за невербальное (подсознательное) мышление. Левое полушарие отвечает за осознание того, что подсознательно посылает ему правое полушарие.
Функциональное состояние структур головного мозга изучается методами регистрации электрических потенциалов. Если регистрирующий электрод располагается в подкорковой структуре, то регистрируемая активность называется субкортикограммой, если в коре мозга - кортикограммой, если электрод располагается на поверхности кожи головы, то через него регистрируется суммарная активность, в которой есть вклад как коры, так и подкорковых структур - это проявление активности называется электроэнцефалограммой (ЭЭГ).
ЭЭГ представляет собой волнообразную кривую, характер которой зависит от состояния коры. Так в покое у человека на ЭЭГ преобладает медленный альфа - ритм (8 -12 Гц, амплитуда = 50 мкВ). При переходе к деятельности идет смена альфа - ритма на быстрый бета - ритм (14 - 30 Гц, амплитуда 25 мкВ). Процесс засыпания сопровождается более медленным тета - ритмом (4 - 7 Гц) или дельта - ритмом (0,5 - 3,5 Гц, амплитуда 100 - 300 мкВ). Когда на фоне покоя или другого состояния мозга человека предъявляется раздражение, например, свет, звук, электрический ток, то с помощью микроэлектродов, вживленных в определенные структуры коры, регистрируются так называемые вызванные потенциалы, латентный период и амплитуда которых зависят от интенсивности раздражения, а компоненты, количество и характер колебаний зависят от адекватности стимула.
Большие полушария головного мозга представляют собой самый массивный отдел головного мозга. Они покрывают мозжечок и ствол мозга. Большие полушария составляют примерно 78% от общей массы мозга. В процессе онтогенетического развития организма большие полушария головного мозга развиваются из коечного мозгового пузыря нервной трубки, поэтому данный отдел головного мозга называется также конечным мозгом.
Большие полушария головного мозга разделены по средней линии глубокой вертикальной щелью на правое и левое полушария.
В глубине средней части оба полушария соединены между собой большой спайкой – мозолистым телом. В каждом полушарии различают доли; лобную, теменную, височную, затылочную и островок.
Доли мозговых полушарий отделяются одна от другой глубокими бороздами. Наиболее важны три глубокие борозды: центральная (роландова) отделяющая лобную долю от теменной, боковая (сильвиева) отделяющая височную долю от теменной, теменно-затылочная отделяющая теменную долю от затылочной на внутренней поверхности полушария.
Каждое полушарие имеет верхнебоковую (выпуклую), нижнюю и внутреннюю поверхность.
Каждая доля полушария имеет мозговые извилины, отделенные друг от друга бороздами. Сверху полушарие покрыто корой ~ тонким слоем серого вещества, которое состоит из нервных клеток.
Кора головного мозга – наиболее молодое в эволюционном отношении образование центральной нервной системы. У человека она достигает наивысшего развития. Кора головного мозга имеет огромное значение в регуляции жизнедеятельности организма, в осуществлении сложных форм поведения и становлении нервно-психических функций.
Под корой находится белое вещество полушарий, оно состоит из отростков нервных клеток – проводников. Из-за образования мозговых извилин общая поверхность коры головного мозга значительно увеличивается. Общая площадь коры полушарий составляет 1200 см2, причем 2/3 ее поверхности находится в глубине борозд, а 1/3 – на видимой поверхности полушарий. Каждая доля мозга имеет различное функциональное значение.
В коре большого мозга выделяют сенсорные, моторные и ассоциативные области.
Сенсорные областиКорковые концы анализаторов имеют свою топографию и на них проецируются определенные афференты проводящих систем. Корковые концы анализаторов разных сенсорных систем перекрываются. Помимо этого, в каждой сенсорной системе коры имеются полисенсорные нейроны, которые реагируют не только на «свой» адекватный стимул, но и на сигналы других сенсорных систем.
Кожная рецептирующая система, таламокортикальные пути проецируются на заднюю центральную извилину. Здесь имеется строгое соматотопическое деление. На верхние отделы этой извилины проецируются рецептивные поля кожи нижних конечностей, на средние - туловища, на нижние отделы - руки, головы.
На заднюю центральную извилину в основном проецируются болевая и температурная чувствительность. В коре теменной доли (поля 5 и 7), где также оканчиваются проводящие пути чувствительности, осуществляется более сложный анализ: локализация раздражения, дискриминация, стереогноз. При повреждениях коры более грубо страдают функции дистальных отделов конечностей, особенно рук.Зрительная система представлена в затылочной доле мозга: поля 17, 18, 19. Центральный зрительный путь заканчивается в поле 17; он информирует о наличии и интенсивности зрительного сигнала. В полях 18 и 19 анализируются цвет, форма, размеры, качества предметов. Поражение поля 19 коры большого мозга приводит к тому, что больной видит, но не узнает предмет (зрительная агнозия, при этом утрачивается также цветовая память).
Слуховая система проецируется в поперечных височных извилинах (извилины Гешля), в глубине задних отделов латеральной (сильвиевой) борозды (поля 41, 42, 52). Именно здесь заканчиваются аксоны задних бугров четверохолмий и латеральных коленчатых тел.Обонятельная система проецируется в области переднего конца гиппокампальной извилины (поле 34). Кора этой области имеет не шести-, а трехслойное строение. При раздражении этой области отмечаются обонятельные галлюцинации, повреждение ее ведет к аносмии (потеря обоняния).Вкусовая система проецируется в гиппокампальной извилине по соседству с обонятельной областью коры.
Моторные области
Впервые Фритч и Гитциг (1870) показали, что раздражение передней центральной извилины мозга (поле 4) вызывает двигательную реакцию. В то же время признано, что двигательная область является анализаторной.В передней центральной извилине зоны, раздражение которых вызывает движение, представлены по соматотопическому типу, но вверх ногами: в верхних отделах извилины - нижние конечности, в нижних - верхние.Спереди от передней центральной извилины лежат премоторные поля 6 и 8. Они организуют не изолированные, а комплексные, координированные, стереотипные движения. Эти поля также обеспечивают регуляцию тонуса гладкой мускулатуры, пластический тонус мышц через подкорковые структуры.В реализации моторных функций принимают участие также вторая лобная извилина, затылочная, верхнетеменная области.Двигательная область коры, как никакая другая, имеет большое количество связей с другими анализаторами, чем, видимо, и обус-ловлено наличие в ней значительного числа полисенсорныхнейронов.
Архитектоника коры больших полушарий мозга
Учение о структурных особенностях строения коры называется архитектоникой. Клетки коры больших полушарий менее специализированы, чем нейроны других отделов мозга; тем не менее определенные их группы анатомически и физиологически тесно связаны с теми или иными специализированными отделами мозга.
Микроскопическое строение коры головного мозга неодинаково в разных ее отделах. Эти морфологические различия коры позволили выделить отдельные корковые цитоархитектонические поля. Имеется несколько вариантов классификаций корковых полей. Большинство исследователей выделяет 50 цитоархитектонических полей, Микроскопическое строение их довольно сложное.
Кора состоит из 6 слоев клеток и их волокон. Основной тип строения коры шестислойной, однако, он не везде однороден. Существуют участки коры, где один из слоев выражен значительно, а другой – слабо. В других областях коры намечается подразделение некоторых слоев на подслои и т.д.
Установлено, что области коры, связанные с определенной функцией, имеют сходное строение. Участки коры, которые близки у животных и человека по своему функциональному значению имеют определенное сходство в строении. Те участки мозга, которые выполняют чисто человеческие функции (речь), имеются только в коре человека, а у животных, даже у обезьян, отсутствуют.
Морфологическая и функциональная неоднородность коры головного мозга позволила выделить центры зрения, слуха, обоняния и т.д., которые имеют свою определенную локализацию. Однако неверно говорить о корковом центре как о строго ограниченной группе нейронов. Специализация участков коры формируется в процессе жизнедеятельности. В раннем детском возрасте функциональные зоны коры перекрывают друг друга, поэтому их границы расплывчаты и нечетки. Только в процессе обучения, накопления собственного опыта практической деятельности происходит постепенная концентрация функциональных зон в отделенные друг от друга центры.Белое вещество больших полушарий состоит из нервных проводников. В соответствии с анатомическими и функциональными особенностями волокна белого вещества делят на ассоциативные, комиссуральные и проекционные. Ассоциативные волокна объединяют различные участки коры внутри одного полушария. Эти волокна бывают короткие и длинные. Короткие волокна обычно имеют дугообразную форму и соединяют соседние извилины. Длинные волокна соединяют отдаленные участки коры. Комиссуальными принято называть те волокна, которые соединяют топографически идентичные участки правого и левого полушарий. Комиссуральные волокна образуют три спайки: переднюю белую спайку, спайку свода, мозолистое тело. Передняя белая спайка соединяет обонятельные области правого и левого полушарий. Спайка свода соединяет между собой гиппокамповые извилины правого и левого полушарий. Основная же масса коммисуальных волокон проходит через мозолистое тело, соединяя между собой симметричные участки обоих полушарий головного мозга.
Проекционными называют те волокна, которые связывают полушария головного мозга с нижележащими отделами мозга – стволом и спинным мозгом. В составе проекционных волокон проходят проводящие пути, несущие афферентную (чувствительную) и эфферентную (двигательную) информацию.
В настоящее время принято делить кору на сенсорные, двигательные, или моторные, и ассоциативные зоны. Такое деление было получено благодаря экспериментам на животных с удалениями различных участков коры, наблюдениями за больными, имеющими патологический очаг в мозге, а также с помощью прямого электрического раздражения коры и периферических структуре регистрацией электрической активности в коре.
В сенсорных зонах представлены корковые концы всех анализаторов. Для зрительного он располагается в затылочной доле мозга (поля 17, 18, 19). В поле 17 заканчивается центральный зрительный путь, информирующий о наличии и интенсивности зрительного сигнала. Поля 18 и 19 анализируют цвет, форму, размеры и качество предмета. При поражении поля 18 больной видит, но не узнает предмета и не различает его цвета (зрительная агнозия).
Корковый конец слухового анализатора локализуется в височной доле коры (извилина Гешля), поля 41, 42, 22. Они участвуют в восприятии и анализе слуховых раздражений, организации слухового контроля речи. Больной, имеющий повреждение поля 22 теряет способность понимать значение произносимых слов.
В височной доле располагается также корковый конец вести булярного анализатора.
Кожный анализатор, а также болевая и температурная чув ствительность проецируются на заднюю центральную извилину, в верхней части которой представлены нижние конечности, в средней - туловище, в нижней - руки и голова.
В коре теменной доли заканчиваются пути соматической чув ствительности, относящиеся к речевой функции, связанной с оценкой воздействия на рецепторы кожи, веса и свойств поверхности, формы и размера предмета.
Корковый конец обонятельного и вкусового анализаторов расположен в гиппокампальной извилине. При раздражении этой области возникают обонятельные галлюцинации, а ее повреждение приводит к аносмии (потере способности ощущать запахи).
Моторные зоны находятся в лобных долях в области передней центральной извилины мозга, раздражение которой вызывает двигательную реакцию. Кора прецентральной извилины (поле 4) представляет первичную двигательную зону. В пятом слое этого поля находятся очень крупные пирамидные клетки (гигантские клетки Беца). Лицо проецируется на нижнюю треть прецентральной извилины, рука занимает ее среднюю треть, туловище и таз - верхнюю треть извилины. Двигательная зона коры для нижних конечностей находится на медиальной поверхности полушария в области передней части парацентральной дольки.
Премоторная область коры (поле 6) располагается кпереди от первичной двигательной зоны. Поле 6 называют вторичной мо торной областью. Ее раздражение вызывает вращение туловища и глаз с подниманием контралатеральной руки. Аналогичные движения наблюдаются у больных во время приступа эпилепсии, если эпилептический очаг локализуется в этой области. Недавно доказана ведущая роль поля 6 в реализации двигательных функций. Поражение поля 6 у человека вызывает резкое ограничение двигательной активности, с трудом выполняются сложные комплексы движений, страдает спонтанная речь.
К полю 6 примыкает поле 8 (лобное глазодвигательное), раздражение которого сопровождается поворотом головы и глаз в сторону, противоположную раздражаемой. Стимуляция различных участков двигательной коры вызывает сокращение соответствующих мышц на противоположной стороне.
Передние отделы лобной коры связывают с «творческим» мышлением. С клинической и функциональной точек зрения интересной областью является нижняя лобная извилина (поле 44). В левом полушарии она связана с организацией двигательных механизмов речи. Раздражение этой области может вызвать вокализацию, но не членораздельную речь, а также прекращение речи, если человек говорил. Поражение этой области приводит к моторной афазии - больной понимает речь, но сам говорить не может.
К ассоциативной коре относят теменно-височно-затылочную, префронтальную и лимбическую области. Она занимает около 80% всей поверхности коры больших полушарий. Ее нейроны обладают мультисенсорными функциями. В ассоциативной коре происходит интеграция различной сенсорной информации и формируется программа целенаправленного поведения, ассоциативная кора окружает каждую проекционную зону, обеспечивая взаимосвязь, например, между сенсорными и моторными областями коры. Нейроны, расположенные в этих областях, обладают полисенсорностью, т.е. способностью отвечать как на сенсорную, так и моторную информацию.
Теменная ассоциативная область коры больших полушарий участвует в формировании субъективного представления об окружающем пространстве, о нашем теле.
Височная область коры участвует в речевой функции посредством слухового контроля речи. При поражении слухового центра речи больной может говорить, правильно излагать свои мысли, но не понимает чужой речи (сенсорная слуховая афазия). Эта область коры играет определенную роль в оценке пространства. Поражение зрительного центра речи приводит к потере способности читать и писать. С височной корой связывают функцию памяти и сновидений.
Лобные ассоциативные поля имеют прямое отношение к лимбическим отделам мозга, они принимают участие в формировании программы сложных поведенческих актов в ответ на воздействие внешней среды на основе сенсорных сигналов всех модальностей.
Особенностью ассоциативной коры является пластичность нейронов, способных к перестройкам в зависимости от поступающей информации. После операции удаления какой-либо области коры в раннем детстве утраченные функции этой области полностью восстанавливаются.
Кора больших полушарий способна, в отличие от нижележащих структур мозга, длительно, в течение всей жизни сохранять следы поступившей информации, т.е. участвовать в механизмах долговременной памяти.
Кора больших полушарий - регулятор вегетативных функций организма («кортиколизация функций»). В ней представлены все безусловные рефлексы, а также внутренние органы. Без коры невозможно выработать условные рефлексы на внутренние органы. При раздражении интерорецепторов методом вызванных потенциалов, электростимуляции и разрушения определенных участков коры доказано ее влияние на деятельность различных органов. Так, разрушение поясной извилины изменяет акт дыхания, функции сердечно-сосудистой системы, желудочно-кишечного тракта. Кора тормозит эмоции - «умейте властвовать собой».
Значение различных участков коры полушарий
головного мозга.
2. Двигательные функции.
3. Функции кожной и проприорицептивной
чувствительности.
4. Слуховые функции.
5. Зрительные функции.
6. Морфологические основы локализации функций в
коре головного мозга.
Ядро двигательного анализатора
Ядро слухового анализатора
Ядро зрительного анализатора
Ядро вкусового анализатора
Ядро кожного анализатора
7. Биоэлектрическая активность головного мозга.
8. Литература.
ЗНАЧЕНИЕ РАЗЛИЧНЫХ УЧАСТКОВ КОРЫ БОЛЬШИХ
ПОЛУШАРИЙ ГОЛОВНОГО МОЗГА
С давних времен между учеными идет спор о местонахождении (локализации) участков коры головного мозга, связанных с различными функциями организма. Были высказаны самые разнообразные и взаимно противоположные точки зрения. Одни считали, что каждой функции нашего организма соответствует строго определенная точка в коре головного мозга, другие отрицали наличие каких бы то ни было центров; любую реакцию они приписывали всей коре, считая ее целиком однозначной в функциональном отношении. Метод условных рефлексов дал возможность И. П. Павлову выяснить ряд неясных вопросов и выработать современную точку зрения.
В коре головного мозга нет строго дробной локализации фун кций. Это следует из экспериментов над животными, когда после разрушения определенных участков коры, например двигательного анализатора, через несколько дней соседние участки берут на себя функцию разрушенного участка и движения животного восстанавливаются.
Эта способность корковых клеток замещать функцию выпавших участков связана с большой пластичностью коры головного мозга.
И. П. Павлов считал, что отдельные области коры имеют разное функциональное значение. Однако между этими областями не существует строго определенных границ. Клетки одной области переходят в соседние области.
Рисунок 1. Схема связи отделов коры с рецепторами.
1 – спинной или продолговатый мозг; 2 – промежуточный мозг; 3 – кора головного мозга
В центре этих областей находятся скопления наиболее специализированных клеток-так называемые ядра анализатора, а на периферии-менее специализированные клетки.
В регуляции функций организма принимают участие не строго очерченные какие-то пункты, а многие нервные элементы коры.
Анализ и синтез поступающих импульсов и формирование ответной реакции на них осуществляются значительно большими областями коры.
Рассмотрим некоторые области, имеющие преимущественно то или иное значение. Схематическое расположение местонахождения этих областей приведено на рисунке 1.
Двигательные функции. Корковый отдел двигательного анализатора расположен главным образом в передней центральной извилине, кпереди от центральной (роландовой) борозды. В этой области находятся нервные клетки, с деятельностью которых связаны все движения организма.
 Отростки
крупных нервных
клеток, находящихся
в глубоких
слоях коры,
спускаются
в продолговатый
мозг, где значительная
часть их перекрещивается,
т. е. переходит
на противоположную
сторону. После
перехода они
опускаются
по спинному
мозгу, где
перекрещивается
остальная
часть. В передних
рогах спинного
мозга они вступают
в контакт с
находящимися
здесь двигательными
нервными клетками.
Таким образом,
возбуждение,
возникшее в
коре, доходит
до двигательных
нейронов передних
рогов спинного
мозга и затем
уже по их волокнам
поступает к
мышцам. Ввиду
того что в
продолговатом,
а частично и
в спинном мозгу
происходит
переход (перекрест)
двигательных
путей на противоположную
сторону, возбуждение,
возникшее в
левом полушарии
головного
мозга, поступает
в правую половину
тела, а в левую
половину тела
поступают
импульсы из
правого полушария.
Вот почему
кровоизлияние,
ранение или
какое-либо
другое поражение
одной из сторон
больших полушарий
влечет за собой
нарушение
двигательной
деятельности
мышц противоположной
половины тела.
Отростки
крупных нервных
клеток, находящихся
в глубоких
слоях коры,
спускаются
в продолговатый
мозг, где значительная
часть их перекрещивается,
т. е. переходит
на противоположную
сторону. После
перехода они
опускаются
по спинному
мозгу, где
перекрещивается
остальная
часть. В передних
рогах спинного
мозга они вступают
в контакт с
находящимися
здесь двигательными
нервными клетками.
Таким образом,
возбуждение,
возникшее в
коре, доходит
до двигательных
нейронов передних
рогов спинного
мозга и затем
уже по их волокнам
поступает к
мышцам. Ввиду
того что в
продолговатом,
а частично и
в спинном мозгу
происходит
переход (перекрест)
двигательных
путей на противоположную
сторону, возбуждение,
возникшее в
левом полушарии
головного
мозга, поступает
в правую половину
тела, а в левую
половину тела
поступают
импульсы из
правого полушария.
Вот почему
кровоизлияние,
ранение или
какое-либо
другое поражение
одной из сторон
больших полушарий
влечет за собой
нарушение
двигательной
деятельности
мышц противоположной
половины тела.
Рисунок 2. Схема отдельных областей коры больших полушарий головного мозга.
1 – двигательная область;
2 – область кожной
и проприорицептивной чувствительности;
3 – зрительная область;
4 – слуховая область;
5 – вкусовая область;
6 – обонятельная область
В передней центральной извилине центры, иннервирующие разные мышечные группы, расположены так, что в верхней части двигательной области находятся центры движений нижних конечностей, затем ниже-центр мышц туловища, еще ниже-центр передних конечностей и, наконец, ниже всех-центры мышц головы.
Центры разных мышечных групп представлены неодинаково и занимают неравномерные области.
Функции кожной и проприоцептивной чувствительности. Область кожной и проприоцептивной чувствительности у человека находится преимущественно позади центральной (роландовой) борозды в задней центральной извилине.
Локализация этой области у человека может быть установлена методом электрического раздражения коры головного мозга во время операций. Раздражение различных участков коры и одновременньш опрос больного об ощущениях, которые он при этом испытывает, дают возможность составить довольно четкое представление об указанной области. С этой же областью связано так называемое мышечное чувство. Импульсы, возникающие в проприорецепторах-рецепторах, находящихся в суставах, сухожилиях н мышцах, поступают преимущественно в этот отдел коры.
Правое полушарие воспринимает импульсы, идущие по центростремительным волокнам преимущественно с левой, а левое полушарие-преимущественно с правой половины тела. Этим объясняется то, что поражение, допустим, правого полушария вызовет нарушение чувствительности преимущественно левой стороны.
Слуховые функции. Слуховая область расположена в височной доле коры. При удалении височных долей нарушаются сложные звуковые восприятия, так как нарушается возможность анализа и синтеза звуковых восприятий.
Зрительные функции. Зрительная область находится в затылочной доле коры головного мозга. При удалении затылочных долей головного мозга у собаки наступает потеря зрения. Животное не видит, натыкается на предметы. Сохраняются только зрачковые рефлексы У человека нарушение зрительной области одного из полушарий вызывает выпадение половины зрения каждого глаза. Если поражение коснулось зрительной области левого полушария, то выпадают функции носовой части сетчатки одного глаза и височной части сетчатки другого глаза.
Такая особенность поражения зрения связана с тем, что зрительные нервы по пути к коре частично перекрещиваются.
Морфологические основы динамической локализации функций в коре полушарий большого мозга (центры мозговой коры).
Знание локализации функций в коре головного мозга имеет огромное теоретическое значение, так как дает представление о нервной регуляции всех процессов организма и приспособлении его к окружающей среде. Оно имеет и большое практическое значение для диагностики мест поражения в полушариях головного мозга.
Представление о локализации функций в коре головного мозга связано прежде всего с понятием о корковом центре. Еще в 1874 г. киевский анатом В. А, Бец выступил с утверждением, что каждый учасгок коры отличается по строению от других участков мозга. Этим было положено начало учению о разнокачественности коры головного мозга - цитоархитектонике (цитос - клетка, архитектонес - строю). В настоящее время удалось выявить более 50 различных участков коры - корковых цитоархитектонических полей, каждое из которых отличается от других по строению и расположению нервных элементов. Из этих полей, обозначаемых номерами, составлена специальная карта мозговой коры человека.
П о
И.П.Павлову,
центр-это мозговой
конец так называемого
анализатора.
Анализатор
- это нервный
механизм, функция
которого состоит
в том, чтобы
разлагать
известную
сложность
внешнего и
внутреннего
мира на отдельные
элементы, т. е.
производить
анализ. Вместе
с тем благодаря
широким связям
с другими
анализаторами
здесь происходит
и синтезирование
анализаторов
друг с другом
и с разными
деятельностями
организма.
о
И.П.Павлову,
центр-это мозговой
конец так называемого
анализатора.
Анализатор
- это нервный
механизм, функция
которого состоит
в том, чтобы
разлагать
известную
сложность
внешнего и
внутреннего
мира на отдельные
элементы, т. е.
производить
анализ. Вместе
с тем благодаря
широким связям
с другими
анализаторами
здесь происходит
и синтезирование
анализаторов
друг с другом
и с разными
деятельностями
организма.
Рисунок 3. Карта цитоархитектонических полей мозга человека (по данным института моэга АМН СССР) Вверху - верхнелатеральная поверхность,внизу- медиальная поверхносгь. Объяснение в тексте.
В настоящее время вся мозговая кора рассматривается как сплошная воспринимающая поверхность. Кора - это совокупность корковых концов анализаторов. С этой точки зрения мы и рассмотрим топографию корковых отделов анализаторов, т. е. главнейшие воспринимающие участки коры полушарий большого мозга.
Прежде всего рассмотрим корковые концы анализаторов, воспринимающих раздражения из внутренней среды организма.
1. Ядро двигательного анализатора, т. е. анализатора проприоцептивных (кинестетических) раздражении, исходящих от костей, суставов, скелетных мышц и их сухожилий, находится в предцентральной извилине (поля 4 и 6} и lobulus paracentralis. Здесь замыкаются двигательные условные рефлексы. Двигательные параличи, возникающие при поражении двигательной зоны, И. П. Павлов объясняет не повреждением двигательных эфферентных нейронов, а нарушением ядра двигательного анализатора, вследствие чего кора не воспринимает кинестетические раздражения и движения становятся невозможными. Клетки ядра двигательного анализатора заложены в средних слоях коры моторной зоны. В глубоких ее слоях (V, отчасти VI) лежат гигантские пирамидные клетки, представляющие собой эфферентные нейроны, которые И. П. Павлов рассматривает как вставочные нейроны, связывающие кору мозга с подкорковыми ядрами, ядрами черепных нервов и передними рогами спинного мозга, т. е. с двигательными нейронами. В предцентральной извилине тело человека, так же как и в задней, спроецировано вниз головой. При этом правая двигательная область связана с левой половиной тела и наоборот, ибо начинающиеся от нее пирамидные пути перекрещиваются частью в продолговатом, а частью в спинном мозге. Мышцы туловища, гортани, глотки находятся под влиянием обоих полушарий. Кроме предцентральной извилины, проприоцептивные импульсы (мышечно-суставная чувствительность) приходят и в кору постцентральной извилины.
2. Ядро двигательного анализатора, имеющего-отношение к сочетанному повороту головы и глаз в противоположную сторону, помещается в средней лобной извилине, в премоторной области (поле 8). Такой поворот происходит и при раздражении поля 17, расположенного в затылочной доле в соседстве с ядром зрительного анализатора. Так как при сокращении мышц глаза в кору мозга (двигательный анализатор, поле 8) всегда поступают не только импульсы от рецепторов этих мышц, но и импульсы от еет-чатки (зрительный анализатор, поле 77), то различные зрительные раздражения всегда сочетаются с различным положением глаз, устанавливаемым сокращением мышц глазного яблока.
3. Ядро двигательного анализатора, посредством которого происходит синтез целенаправленных сложных профессиональных, трудовых и спортивных движений, помещается в левой (у правшей) нижней теменной дольке, в gyrus supramarginalis (глубокие слои поля 40). Эти координированные движения, образованные по принципу временных связей и выработанные практикой индивидуальной жизни, осуществляются через связь gyrus supramarginalis с предцентральной извилиной. При поражении поля 40 сохраняется способность к движению вообще, но появляется неспособность совершать целенаправленные движения, действовать - апраксия (праксия - действие, практика).
4. Ядро анализатора положения и движения головы - статический анализатор (вестибулярный аппарат) в коре мозга точно еще не локализован. Есть основания предполагать, что вестибулярный аппарат проецируется в той же области коры, что и улитка, т. е. в височной доле. Так, при поражении полей 21 и 20, лежащих в области средней и нижней височных извилин, наблюдается атаксия, т. е расстройство равновесия, покачивание тела при стоянии. Этот анализатор, играющий решающую роль в прямохождении человека, имеет особенное значение для работы летчиков в условиях реактивной авиации, так как чувствительность вестибулярного аппарата на самолете значительно понижается.
5. Ядро анализатора импульсов, идущих от внутренностей и сосудов, находится в нижних отделах передней и задней центральных извилин. Центростремительные импульсы от внутренностей, сосудов, непроизвольной мускулатуры и желез кожи поступают в этот отдел коры, откуда отходят центробежные пути к подкорковым вегетативным центрам.
В премоторной области (поля 6 и 8) совершается объединение вегетативных функций.
Нервные импульсы из внешней среды организма поступают в корковые концы анализаторов внешнего мира.
1. Ядро слухового анализатора лежит в средней части верхней височной извилины, на поверхности, обращенной к островку, - поля 41, 42, 52, где проецирована улитка. Повреждение ведет к глухоте.
2. Ядро зрительного анализатора находится в затылочной доле - поля 18, 19. На внутренней поверхности затылочной доли, по краям sulcus Icarmus, в поле 77 заканчивается зрительный путь. Здесь спроецирована сетчатка глаза. При поражении ядра зрительного анализатора наступает слепотa. Выше поля 17 расположено поле 18, при поражении которого зрение сохраняется и только теряется зрительная память. Еще выше находится поле при поражении которого утрачивается ориентация в непривычной обстанвке.
3. Ядро вкусового анализатора, по одним данным, находится в нижней постцентральной извилине, близко к центрам мышц рта и языка, по другим - в ближайшем соседстве с корковым концом обонятельного анализатора, чем объясняется тесная связь обонятельных и вкусовых ощу-ний. Установлено, что расстройство вкуса наступает при поражении поля 43.
Анализаторы обоняния, вкуса и слуха каждого полушария связаны с рецепторами соответствующих органов обеих сторон тела.
4. Ядро кожного анализатора (осязательная, болевая и температурная чувствительность) находится в постцентральной извилине (поля 7, 2, 3) и в пе верхней теменной области (поля 5 и 7).
Частный вид кожной чувствительности - узнавание предметов на ощупь - стереогнозия (стереос - пространственный, гнозис - знание) связана с участком коры верхней теменной дольки (поле 7) перекрестно: левое полушарие соответствует правой руке, правое - левой руке. При поражении поверхностных слоев поля 7 утрачивается способность узнавать предметы на ощупь, при закрытых глазах.
Биоэлектрическая активность головного мозга.
Отведение биопотенциалов головного мозга - электроэнцефалография-дает представление об уровне физиологической активности головного мозга. Кроме метода электроэнцефалографии-записи биоэлектрических потенциалов, используется метод энцефалоскопии-регистрации колебаний яркости свечения множества точек мозга (от 50 до 200).
Электроэнцефалограмма является интегративным пространственно-временным показателем спонтанной электрической активности мозга. В ней различают амплитуду (размах) колебаний в микровольтах и частоту колебаний в герцах. В соответствии с этим в электроэнцефалограмме различают четыре типа волн: -, -, - и -ритмы. Для -ритма характерны частоты в диапазоне 8-15 Гц, при амплитуде колебаний 50-100 мкВ. Он регистрируется только у людей и высших обезьян в состоянии бодрствования, при закрытых глазах и при отсутствии внешних раздражителей. Зрительные раздражители тормозят -ритм.
У отдельных людей, обладающих живым зрительным воображением, -ритм может вообще отсутствовать.
Для деятельного мозга характерен (-ритм. Это электрические волны с амплитудой от 5 до 30 мкВ и частотой от 15 до 100 Гц Он хорошо регистрируется в лобных и центральных областях головного мозга. Во время сна появляется -ритм. Он же наблюдается при отрицательных эмоциях, болезненных состояниях. Частота потенциалов -ритма от 4 до 8 Гц, амплитуда от 100 до 150 мкВ Во время сна появляется и -ритм - медленные (с частотой 0,5-3,5 Гц), высокоамплитудные (до 300 мкВ) колебания электрической активности мозга.
Помимо рассмотренных видов электрической активности, у человека регистрируется Е-волна (волна ожидания раздражителя) и веретенообразные ритмы. Волна ожидания регистрируется при выполнении сознательных, ожидаемых действий. Она предшествует появлению ожидаемого раздражителя во всех случаях, даже при неоднократном его повторении. По-видимому, ее можно рассматривать как электроэнцефалографический коррелят акцептора действия, обеспечивающего предвидение результатов действия до его завершения. Субъективная готовность отвечать на действие стимула строго определенным образом достигается психологической установкой (Д. Н. Узнадзе). Веретенообразные ритмы непостоянной амплитуды, с частотой от 14 до 22 Гц, появляются во время сна. Различные формы жизне деятельности приводят к существенному изменению ритмов биоэлектрической активности мозга.
При умственной работе усиливается -ритм, -ритм при этом исчезает. При мышечной работе статического характера наблюдается десинхронизация электрической активности мозга. Появляются быстрые колебания с низкой амплитудой.Во время динамической работы пе-. риоды десинхронизированной и синхронизированной активности наблюдаются соответственно в моменты рабогы и отдыха.
Образование условного рефлекса сопровождается десинхронизацией волновой активности мозга.
Десинхронизация волн происходит при переходе от сна к бодрствованию. При этом веретенообразные ритмы сна сменяются
-ритмом, увеличивается электрическая активность ретикулярной формации. Синхронизация (одинаковые по фазе и направлению волны)
характерна для тормозного процесса. Она наиболее отчетливо выражена при выключении ретикулярной формации стволовой части мозга. Волны электроэнцефалограммы, по мнению большинства исследователей, являются результатом суммации тормозных и возбуждающих постсинаптических потенциалов. Электрическая активность мозга не является простым отражением обменных процессов в нервной ткани. Установлено, в частности, что в импульсной активности отдельных скоплений нервных клеток обнаруживаются признаки акустического и семантического кодов.
Кроме специфических ядер таламуса возникают и развиваются ассоциативные ядра, имеющие связи с неокортексом и определяющие развитие конечного мозга. Третьим источником афферентных воздействий на кору больших полушарий является гипоталамус, который играет роль высшего регуляторного центра вегетативных функций. У млекопитающих филогенетически более древние отделы переднего гипоталамуса связаны с...
Затрудняется формирование условных рефлексов, нарушаются процессы памяти, теряется избирательность реакций и отмечается неумеренное их усиление. Большой мозг состоит из почти идентичных половин – правого и левого полушарий, которые связаны мозолистым телом. Комиссуральные волокна связывают симметричные зоны коры. Тем не менее, кора правого и левого полушарий не симметричны не только внешне, но и...
Подход к оценке механизмов работы высших отделов головного мозга с использованием условных рефлексов был столь успешным, что позволил Павлову создать новый раздел физиологии - «Физиологию высшей нервной деятельности», науку о механизмах работы больших полушарий головного мозга. БЕЗУСЛОВНЫЕ И УСЛОВНЫЕ РЕФЛЕКСЫ Поведение животных и человека представляет собой сложную систему взаимосвязанных...