Yağ turşularının oksidləşməsi (β-oksidləşmə). sonrakı ardıcıllıq. reaksiyalar. B-yağ turşularının oksidləşməsi Palmitik turşunun oksidləşməsi
Artıq qeyd edildiyi kimi, heyvan orqanizmi oksidləşmə prosesində çıxarılan enerjinin əhəmiyyətli hissəsini β-karbon atomunda oksidləşmə ilə parçalanan yağ turşularından alır.
Yağ turşularının β-oksidləşməsi ilk dəfə 19004-cü ildə F.Knoop tərəfindən tədqiq edilmişdir. Sonradan məlum oldu ki, β-oksidləşmə yalnız mitoxondriyada baş verir. F. Linen və onun həmkarlarının (1954-1958) işi sayəsində yağ turşularının oksidləşməsinin əsas enzimatik prosesləri aydınlaşdırıldı. Yağ turşularının oksidləşməsinin bu yolunu kəşf edən alimlərin şərəfinə β-oksidləşmə prosesi adlanır. Knoop-Kətan dövrü.
β-oksidləşmə- asetil-KoA şəklində yağ turşusunun karboksil ucundan ardıcıl olaraq 2 karbon atomunun ayrıldığı yağ turşusu katabolizminin xüsusi yolu. Metabolik yol - β-oksidləşmə - belə adlandırılmışdır, çünki yağ turşularının oksidləşmə reaksiyaları β-karbon atomunda baş verir. β-oksidləşmə və asetil-KoA-nın TCA dövründə (trikarboksilik turşu dövrü) sonrakı oksidləşməsi reaksiyaları oksidləşdirici fosforlaşma mexanizmi vasitəsilə ATP sintezi üçün əsas enerji mənbələrindən biri kimi xidmət edir. Yağ turşularının β-oksidləşməsi yalnız aerob şəraitdə baş verir.
Bütün çoxmərhələli oksidləşmə reaksiyaları xüsusi fermentlər tərəfindən sürətləndirilir. Yüksək yağ turşularının β-oksidləşməsi bütün canlı orqanizmlərdə baş verən universal biokimyəvi prosesdir. Məməlilərdə bu proses bir çox toxumalarda, xüsusilə qaraciyər, böyrəklər və ürəkdə baş verir. Yağ turşularının oksidləşməsi mitoxondriyada baş verir. Doymamış yüksək yağ turşuları (oleik, linoleik, linolenik və s.) ilkin olaraq doymuş turşulara qədər azaldılır.
Yağ turşularının mitoxondrial matrisə nüfuz etməsindən əvvəl onların aktivləşdirmə ilə əlaqə yaratmaqla koenzim A(HS~CoA), yüksək enerjili bağ ehtiva edir. Sonuncu, görünür, meydana gələn birləşmənin oksidləşmə reaksiyalarının daha hamar bir gedişinə kömək edir. asil koenzim A(asil-CoA).
Yüksək yağ turşularının CoA ilə qarşılıqlı əlaqəsi xüsusi ligazalarla sürətlənir - asil-CoA sintetazalarıüç növ, müvafiq olaraq qısa, orta və uzun karbohidrogen radikalları olan turşular üçün xüsusidir. Onlar endoplazmatik retikulumun membranlarında və mitoxondriyanın xarici membranında lokallaşdırılmışdır. Bütün asil-CoA sintetazaları multimerlər kimi görünür; Beləliklə, qaraciyər mikrosomlarından olan ferment 168 kDa molekulyar çəkiyə malikdir və 6 eyni subunitdən ibarətdir. Yağ turşularının aktivləşmə reaksiyası 2 mərhələdə baş verir:
a) birincisi, yağ turşusu ATP ilə reaksiyaya girərək asiladenilat əmələ gətirir:
RCOOH + ATP → RCO~AMP + FF
b) sonra asil-CoA-nın aktivləşdirilmiş formasının əmələ gəlməsi baş verir:
RCO~AMF + NS~KoA → RCO~SKoA + AMF
Pirofosfat (PP) pirofosfataza tərəfindən sürətlə hidrolizə olunur, bunun nəticəsində bütün reaksiya geri dönməzdir: PP + H 2 O → 2P
Xülasə tənliyi:
RCOOH + ATP+ HS~CoA→ RCO~SKoA + AMF + 2P
Qısa və orta zəncir uzunluğuna malik yağ turşuları (4-dən 12 karbon atomuna qədər) mitoxondrial matrisə diffuziya yolu ilə nüfuz edə bilər, burada onların aktivləşməsi baş verir. İnsan orqanizmində üstünlük təşkil edən uzun zəncirli yağ turşuları (12-20 karbon atomu) mitoxondriyanın xarici membranında yerləşən asil-CoA sintetazaları tərəfindən aktivləşdirilir.
Daxili mitoxondrial membran sitoplazmada əmələ gələn uzun zəncirli asil-CoA-ları keçirməzdir. Aktivləşdirilmiş yağ turşularının daşıyıcısı kimi xidmət edir karnitin (vitamin B t), qidadan gəlir və ya əsas amin turşuları lizin və metionindən sintez olunur.
Mitoxondriyanın xarici membranı var karnitin asiltransferaza I fermenti(karnitin palmitoiltransferaza I), asilkarnitinin əmələ gəlməsi ilə reaksiyanı kataliz edir:
RCO~SKoA + H 3 C- N + -CH 2 -CH-CH 2 -COOH ↔ H 3 C- N + -CH 2 -CH-CH 2 -COOH + HS~KoA
Acyl-CoA Carnitine (B t) Acylcarnitine Coenzyme A
Bu ferment tənzimləyicidir, asil qruplarının mitoxondriyaya daxil olma sürətini və nəticədə yağ turşularının oksidləşmə sürətini tənzimləyir.
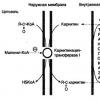
Yaranan asilkarnitin membranlararası boşluqdan daxili membranın xarici tərəfinə keçir və karnitin asilkarnitin translokazı ilə daxili mitoxondrial membranın daxili səthinə nəql olunur, burada ferment karnitin asiltransferaza II asilin intramitoxondrial CoA-ya köçürülməsini, yəni əks reaksiyanı katalizləyir (şək. 9).

Şəkil 9. Uzun karbohidrogen radikalları olan yağ turşularının mitoxondrial membranlar vasitəsilə ötürülməsi
Beləliklə, asil-CoA β-oksidləşmə fermentləri üçün əlçatan olur. Sərbəst karnitin eyni translokazla daxili mitoxondrial membranın sitozolik tərəfinə qaytarılır. Bundan sonra asil-CoA β-oksidləşmə reaksiyalarına daxil olur.
Mitoxondrial matrisdə asil-CoA-nın katabolizmi (parçalanması) təkrarlanan ardıcıllıq nəticəsində baş verir. dörd reaksiya.
1) Hər bir dövrədə ilk reaksiya onun ferment tərəfindən oksidləşməsidir asil-CoA dehidrogenaz, koenzimi FAD olan. Dehidrogenləşmə β və α karbon atomları arasında baş verir, nəticədə karbon zəncirində ikiqat bağ əmələ gəlir və bu reaksiyanın məhsulu enoil-CoA olur:
R-CH 2 -CH 2 CO~SKoA + FAD → R-CH=CHCO~SkoA + FADN 2
Acyl-CoA Enoil-CoA
2) Yağ turşusunun oksidləşmə dövrünün ikinci pilləsində enol-CoA ikiqat bağı hidratlanır, nəticədə β-hidroksiasil-CoA əmələ gəlir. Reaksiya bir ferment tərəfindən katalizlənir enol-CoA hidrataz:
R-CH=CHCO~SKoA +H 2 O → R-CH-CH 2 CO~SKoA
Enoil-CoA β-hidroksiasil-CoA
3) Dövrün üçüncü mərhələsində β-hidroksiasil-CoA fermentin iştirakı ilə dehidrogenləşməyə (ikinci oksidləşmə) məruz qalır. β-hidroksiasil-CoA dehidrogenaz, koenzimi NAD+ olan. Bu reaksiyanın məhsulu β-ketoasil-CoA-dır:
R-CH-CH 2 CO~SKoA + NAD + → R-COCH 2 CO~SkoA + NADH + H +
β-hidroksiasil-CoA β-ketoasil-CoA
4) Yağ turşularının oksidləşmə dövrünün son reaksiyası kataliz edilir asetil-KoA asiltransferaza (tiolaza). Bu mərhələdə β-ketoasil-KoA sərbəst CoA ilə reaksiyaya girir və ilk növbədə asetil-KoA şəklində ana yağ turşusunun iki terminal karbon atomunu ehtiva edən iki karbonlu fraqment, ikincisi isə CoA əmələ gətirmək üçün parçalanır. yağ turşusu esteri, indi iki karbon atomu ilə qısaldılmışdır. Hidrolizlə analoji olaraq bu reaksiya deyilir tioliz:
R-COCH 2 CO~SKoA + HS~KoA → CH 3 CO~SkoA + R 1 CO~SkoA
β-ketoasil-CoA Asetil-CoA Asil-CoA,
tərəfindən qısaldılmışdır
2 karbon atomu
Qısaldılmış asil-CoA daha sonra asil-CoA dehidrogenaz (oksidləşmə) tərəfindən katalizləşdirilmiş reaksiya ilə başlayan növbəti oksidləşmə dövrünə keçir, ardınca hidratasiya reaksiyası, ikinci oksidləşmə reaksiyası, tiolaza reaksiyası, yəni bu proses dəfələrlə təkrarlanır. (şək. 10).
β- Yüksək yağ turşularının oksidləşməsi mitoxondriyada baş verir. Tənəffüs dövrünün fermentləri də onlarda lokallaşdırılmışdır ki, bu da hidrogen atomlarının və elektronların ADP-nin oksidləşdirici fosforlaşması şəraitində oksigenə keçməsinə səbəb olur, buna görə də yüksək yağ turşularının β-oksidləşməsi ATP sintezi üçün enerji mənbəyidir.

Şəkil 10. Yağ turşularının oksidləşməsi
Yüksək yağ turşularının β-oksidləşməsinin son məhsulu karbon atomlarının cüt sayı edir asetil-KoA, A qəribə ilə- propionil-CoA.
Əgər asetil-KoA bədəndə yığılsa, onda HS~KoA ehtiyatları tezliklə tükənəcək və yüksək yağ turşularının oksidləşməsi dayanacaq. Lakin bu baş vermir, çünki CoA asetil-KoA-dan tez ayrılır. Bir sıra proseslər buna gətirib çıxarır: asetil-KoA trikarboksilik və dikarboksilik turşuların dövrünə və ya ona çox yaxın olan qlioksil dövrünə daxil edilir və ya asetil-KoA sterolların və izoprenoid qrupları olan birləşmələrin sintezi üçün istifadə olunur, və s.
Propionil-KoA, tək sayda karbon atomu olan yüksək yağ turşularının β-oksidləşməsinin son məhsulu olan süksinil-KoA-ya çevrilir və trikarboksilik və dikarboksilik turşuların dövrü ilə istifadə olunur.
İnsan bədənindəki yağ turşularının təxminən yarısı doymamış .
Bu turşuların β-oksidləşməsi adi şəkildə ikiqat bağ üçüncü və dördüncü karbon atomları arasında olana qədər davam edir. Sonra ferment enol-CoA izomeraza qoşa bağı 3-4-cü mövqedən 2-3-cü mövqeyə keçirir və qoşa bağın cis-ini β-oksidləşmə üçün lazım olan trans konformasiyaya dəyişir. Bu β-oksidləşmə dövründə ilk dehidrogenləşmə reaksiyası baş vermir, çünki yağ turşusu radikalında ikiqat bağ artıq mövcuddur. Bundan əlavə, β-oksidləşmə dövrləri davam edir, adi yoldan fərqlənmir. Yağ turşularının mübadiləsinin əsas yolları Şəkil 11-də göstərilmişdir.

Şəkil 11. Yağ turşularının mübadiləsinin əsas yolları
Bu yaxınlarda aşkar edilmişdir ki, β-oksidləşmə ilə yanaşı, yağ turşularının katabolizmasının əsas yolu, beyin toxuması yağ turşularının α-oksidləşməsi karbon atomlarının sayı ilə (C 13 -C 18), yəni molekulun karboksil ucundan bir karbonlu fraqmentlərin ardıcıl olaraq aradan qaldırılması.
Bu tip oksidləşmə ən çox bitki toxumalarında olur, lakin bəzi heyvan toxumalarında da baş verə bilər. α-Oksidləşmə tsiklik xarakter daşıyır və dövr iki reaksiyadan ibarətdir.
Birinci reaksiya yağ turşusunun hidrogen peroksid ilə müvafiq aldehid və CO 2-ə xüsusi bir maddənin iştirakı ilə oksidləşməsindən ibarətdir. peroksidazlar:

Bu reaksiya nəticəsində karbohidrogen zənciri bir karbon atomu ilə qısalır.
İkinci reaksiyanın mahiyyəti, əmələ gələn aldehidin təsiri altında müvafiq karboksilik turşuya nəmləndirilməsi və oksidləşməsidir. aldehid dehidrogenaz NAD koenziminin oksidləşmiş formasını ehtiva edir:
α-oksidləşmə dövrü yenidən təkrarlanır. β-oksidləşmə ilə müqayisədə bu tip oksidləşmə enerji baxımından daha az əlverişlidir.
ω-yağ turşularının oksidləşməsi. Heyvanların və bəzi mikroorqanizmlərin qaraciyərində yağ turşularının ω-oksidləşməsini, yəni ω hərfi ilə təyin olunan terminal CH 3 qrupunda oksidləşməni təmin edən ferment sistemi mövcuddur. Əvvəlcə təsir altında monooksigenazlar hidroksilləşmə ω-hidroksi turşusu əmələ gətirir:

Daha sonra ω-hidroksi turşusu müvafiq maddələrin təsiri ilə ω-dikarboksilik turşuya oksidləşir. dehidrogenazlar:
Beləliklə alınan ω-dikarboksilik turşu β-oksidləşmə reaksiyaları ilə hər iki ucunda qısaldılır.
Yağ turşularının tərkibindəki enerjini ATP bağlarının enerjisinə çevirmək üçün yağ turşularının CO 2 və suya oksidləşməsinin metabolik yolu var ki, bu da trikarboksilik turşu dövrü və tənəffüs zənciri ilə sıx bağlıdır. Bu yol adlanır β-oksidləşmə, çünki yağ turşusunun 3-cü karbon atomunun (β-mövqeyi) karboksil qrupuna oksidləşməsi baş verir və eyni zamanda ilkin yağ turşusunun C 1 və C 2 də daxil olmaqla asetil qrupu turşudan ayrılır.
β-oksidləşmənin elementar diaqramı
β-oksidləşmə reaksiyaları baş verir mitoxondriya bədənin əksər hüceyrələri (sinir hüceyrələrindən başqa). Oksidləşmə üçün qandan sitozola daxil olan və ya öz hüceyrədaxili TAG-larının lipolizi zamanı meydana çıxan yağ turşuları istifadə olunur. Palmitik turşunun oksidləşməsinin ümumi tənliyi aşağıdakı kimidir:
Palmitoyl-ScoA + 7FAD + 7NAD + + 7H 2 O + 7HS-KoA → 8Asetil-SCoA + 7FADH 2 + 7NADH
Yağ turşularının oksidləşməsinin mərhələləri
1. Mitoxondrial matriksə nüfuz etmədən və oksidləşmədən əvvəl yağ turşusu aktivləşdirin sitozolda. Bu, asil-SCoA yaratmaq üçün ona koenzim A əlavə etməklə həyata keçirilir. Acyl-ScoA yüksək enerjili birləşmədir. Reaksiyanın dönməzliyi difosfatın iki fosfor turşusuna hidrolizi ilə əldə edilir.
Acyl-ScoA sintetazaları endoplazmatik retikulumda, mitoxondrilərin xarici membranında və onların içərisində olur. Müxtəlif yağ turşuları üçün spesifik sintetazaların geniş spektri mövcuddur.
Yağ turşularının aktivləşdirilməsi reaksiyası
2. Acyl-ScoA mitoxondrial membrandan keçə bilmir, buna görə də onu vitaminə bənzər bir maddə karnitin (vitamin B11) ilə birlikdə köçürməyin bir yolu var. Mitoxondriyanın xarici membranında bir ferment var karnitin asiltransferaza I.
![]()
Karnitindən asılı olaraq yağ turşularının mitoxondriyə daşınması
Karnitin qaraciyər və böyrəklərdə sintez olunur və sonra digər orqanlara daşınır. In intrauterin dövr və erkən illər Həyatda karnitinin orqanizm üçün əhəmiyyəti son dərəcə böyükdür. Sinir sisteminə enerji təchizatı uşaq bədən və xüsusilə beyin iki paralel proses sayəsində həyata keçirilir: yağ turşularının karnitindən asılı oksidləşməsi və qlükozanın aerob oksidləşməsi. Karnitin beyin və onurğa beyninin böyüməsi, hərəkət və əzələlərin qarşılıqlı təsirindən məsul olan sinir sisteminin bütün hissələrinin qarşılıqlı əlaqəsi üçün lazımdır. Karnitin çatışmazlığını əlaqələndirən tədqiqatlar var serebral iflic və fenomen" beşikdə ölüm".
Körpələr, vaxtından əvvəl doğulmuş körpələr və az çəkisi olan körpələr karnitin çatışmazlığına xüsusilə həssasdırlar. Onların endogen ehtiyatları müxtəlif stresli vəziyyətlərdə (infeksion xəstəliklər, mədə-bağırsaq traktının pozğunluqları, qidalanma pozğunluqları) tez tükənir. Karnitin biosintezi qeyri-kafidir və adi qidalardan qəbul qanda və toxumalarda kifayət qədər səviyyəni saxlaya bilmir.
3. Karnitinə bağlandıqdan sonra yağ turşusu translokaz vasitəsilə membrandan keçir. Burada, membranın daxili tərəfində, karnitin asiltransferaza II fermenti yenidən β-oksidləşmə yoluna daxil olan asil-SCoA əmələ gətirir.
4. Prosesin özü β-oksidləşmə tsiklik təkrarlanan 4 reaksiyadan ibarətdir. Onlar ardıcıl olaraq baş verir oksidləşmə(asil-ScoA dehidrogenaz), nəmləndirmə(enoyl-SCoA hidrataz) və yenidən oksidləşmə 3-cü karbon atomu (hidroksiasil-ScoA dehidrogenaz). Sonuncu transferaz reaksiyasında asetil-SCoA yağ turşusundan ayrılır. Qalan (iki karbonla qısaldılmış) yağ turşusuna HS-CoA əlavə edilir və o, birinci reaksiyaya qayıdır. Bu, son dövrədə iki asetil-SCoA əmələ gələnə qədər təkrarlanır.

Yağ turşularının β-oksidləşməsi reaksiyalarının ardıcıllığı
β-oksidləşmənin enerji balansının hesablanması
Əvvəllər oksidləşmə səmərəliliyini hesablayarkən NADH üçün P/O əmsalı 3.0, FADH 2 - 2.0-a bərabər alınırdı.
Müasir məlumatlara görə, NADH üçün P/O əmsalının dəyəri 2,5, FADH 2 - 1,5-ə uyğundur.
Yağ turşularının β-oksidləşməsi zamanı əmələ gələn ATP miqdarını hesablayarkən nəzərə almaq lazımdır:
- əmələ gələn asetil-SCoA miqdarı yağ turşusunda karbon atomlarının sayının adi olaraq 2-yə bölünməsi ilə müəyyən edilir.
- nömrə β-oksidləşmə dövrləri. β-oksidləşmə dövrlərinin sayını iki karbon vahidlərinin zənciri kimi yağ turşusu konsepsiyasına əsaslanaraq müəyyən etmək asandır. Bölmələr arasındakı fasilələrin sayı β-oksidləşmə dövrlərinin sayına uyğundur. Eyni dəyəri (n/2 -1) düsturundan istifadə etməklə hesablamaq olar, burada n turşudakı karbon atomlarının sayıdır.
- yağ turşusunda ikiqat bağların sayı. Birinci β-oksidləşmə reaksiyasında FAD-ın iştirakı ilə ikiqat bağ əmələ gəlir. Əgər yağ turşusunda artıq qoşa bağ varsa, o zaman bu reaksiyaya ehtiyac yoxdur və FADN 2 əmələ gəlmir. İtirilmiş FADN 2-nin sayı ikiqat istiqrazların sayına uyğundur. Dövrün qalan reaksiyaları dəyişmədən davam edir.
- aktivləşdirməyə sərf olunan ATP enerjisinin miqdarı (həmişə iki yüksək enerji bağına uyğundur).
Misal. Palmitik turşunun oksidləşməsi
- 16 karbon atomu olduğu üçün β-oksidləşmə əmələ gəlir 8 asetil-SCoA molekulu. Sonuncu TCA dövrünə daxil olur, dövrün bir növbəsində oksidləşdikdə 3 molekul NADH (7,5 ATP), 1 molekul FADH 2 (1,5 ATP) və 1 molekul GTP əmələ gəlir ki, bu da 10 molekula bərabərdir. ATP. Beləliklə, 8 molekul asetil-SCoA 8 × 10 = əmələ gəlməsini təmin edəcəkdir. 80 ATP molekulları.
- Palmitik turşu üçün β-oksidləşmə dövrlərinin sayı 7-dir. Hər dövrədə 1 molekul FADH 2 (1,5 ATP) və 1 molekul NADH (2,5 ATP) istehsal olunur. Tənəffüs zəncirinə girərək, ümumilikdə 4 ATP molekulunu "verirlər". Beləliklə, 7 dövrədə 7 × 4 = 28 ATP molekulu əmələ gəlir.
- Palmitik turşuda ikiqat bağlar Yox.
- Yağ turşusunu aktivləşdirmək üçün 1 molekul ATP istifadə olunur, lakin o, AMP-yə hidrolizə olunur, yəni israf olunur. 2 makroergik əlaqə və ya iki ATP.
- Beləliklə, yekunlaşdıraraq, əldə edirik 80+28-2 =106 Palmitik turşunun oksidləşməsi zamanı ATP molekulları əmələ gəlir.
Karbohidratlar insan pəhrizinin əsas hissəsini təşkil edir və bədənin enerji ehtiyacının əhəmiyyətli bir hissəsini təmin edir. Balanslaşdırılmış pəhriz ilə gündəlik karbohidrat miqdarı zülal və yağların miqdarından orta hesabla 4 dəfə çoxdur.
Qidalanmada karbohidratların rolu:
1. Karbohidratlar edir enerji funksiyası. 1 q karbohidrat oksidləşdikdə 4,1 kkal enerji ayrılır. Karbohidratların əsas hissəsinin parçalandığı qlükoza bədəndəki əsas enerji substratıdır.
2. Əzələ fəaliyyətiəhəmiyyətli qlükoza istehlakı ilə müşayiət olunur. Fiziki iş zamanı ilk növbədə karbohidratlar istehlak edilir və yalnız onların ehtiyatları (qlikogen) tükəndikdə mübadiləə yağlar daxil edilir.
3. Karbohidratlar normal fəaliyyət üçün vacibdir Mərkəzi sinir sistemi, hüceyrələri qanda qlükoza çatışmazlığına çox həssasdır.
4. Karbohidratlar edir struktur funksiyası. Sadə karbohidratlar birləşdirici toxumanın əsasını təşkil edən qlikoproteinlərin əmələ gəlməsi mənbəyi kimi xidmət edir.
5. Karbohidratlar iştirak edir zülalların və yağların metabolizmasında. Yağlar karbohidratlardan əmələ gələ bilər.
6. Bitki mənşəli karbohidratlar (selüloz, pektin maddələri) bağırsaq hərəkətliliyini stimullaşdırır və orada yığılan zəhərli məhsulların xaric olmasına kömək edir.
Mənbələr karbohidratlar üstünlük təşkil edir bitki məhsulları, xüsusilə un məmulatları, taxıllar, şirniyyatlar. Əksər qidalarda karbohidratlar nişasta şəklində və daha az dərəcədə disaxaridlər (süd, şəkər çuğunduru, meyvə və giləmeyvə) şəklində təqdim olunur. Karbohidratların daha yaxşı mənimsənilməsi üçün onların əksəriyyətinin bədənə nişasta şəklində daxil olması lazımdır.
Nişasta mədə-bağırsaq traktında tədricən qlükozaya parçalanır, qana kiçik hissələrlə daxil olur, bu da onun istifadəsini yaxşılaşdırır və qan şəkərinin sabit səviyyəsini saxlayır. Böyük miqdarda şəkər bir anda tətbiq edildikdə, qanda qlükozanın konsentrasiyası kəskin şəkildə artır və sidiklə xaric olunmağa başlayır. Ən əlverişli şərtlər karbohidratların 64% nişasta, 36% isə şəkər şəklində istehlak edildikdə hesab olunur.
İstehlak dərəcəsi karbohidratlar işin intensivliyindən asılıdır. Fiziki iş zamanı karbohidratlar daha böyük miqdarda tələb olunur. Orta hesabla 1 kq bədən çəkisi üçün tələb olunur 4-6-8 gündə g karbohidratlar, yəni. zülal və yağlardan təxminən 4 dəfə çoxdur.
Həddindən artıq karbohidrat qəbulu piylənməyə və mədə-bağırsaq traktının həddindən artıq yüklənməsinə səbəb ola bilər, çünki karbohidratlarla zəngin olan bitki qidaları adətən daha həcmlidir, ağırlıq hissi yaradır və yeməyin ümumi həzmini pozur.
Karbohidratların olmaması hipoqlikemik şərtlərin inkişaf riski səbəbindən qidada da arzuolunmazdır. Karbohidrat çatışmazlığı, bir qayda olaraq, ümumi zəiflik, yuxululuq, yaddaşın azalması, zehni və fiziki performans, baş ağrısı, zülalların, vitaminlərin həzm qabiliyyətinin azalması, asidoz və s. ilə müşayiət olunur.Bu baxımdan, gündəlik qida rasionunda karbohidratların miqdarı olmamalıdır 300 q-dan az olmalıdır
Karbohidratlar qrupuna yaxından insan orqanizmi tərəfindən zəif həzm olunan əksər bitki qidalarında olan maddələr - pektin maddələri (həzm olunmayan karbohidratlar) və lif daxildir.
Pektik maddələrdir yüksək sorbsiya (udma) qabiliyyətinə malik olan bitki jeli maddələri. Onlar həzm sistemi xəstəliklərinin, yanıqların və xoraların müalicəsində faydalı təsir göstərir, həmçinin bəzi zəhərli maddələri zərərsizləşdirmək qabiliyyətinə malikdirlər (onlar xüsusilə ağır metal duzlarının, məsələn, qurğuşun birləşmələrinin orqanizmdən çıxarılmasında fəaldırlar).
Portağal, alma, qara qarağat və digər meyvə və giləmeyvələrdə çoxlu pektin maddəsi var.
Sellüloza(digər adlar - qaba tərəvəz və ya həzm olunmayan və ya qida və ya pəhriz lifi) bitki qidalarının kütləvi hüceyrə divarlarının bir hissəsi olan bir polisaxariddir. Lifli, kifayət qədər qaba bir quruluşa malikdir.
Pəhriz lifinin ümumi mənbələri kəpək, çörək və dənli bitkilərdir (xüsusilə qarabaşaq yarması və yulaf ezmesi). Böyük miqdarda bir çox tərəvəz, meyvə, yarpaq və bitkilərin gövdələrində olur; xüsusilə taxılların qabıqlarında və meyvələrin qabıqlarında çox olur. Tərəvəz və meyvələri konservləşdirərkən pəhriz lifi tamamilə qorunur (pulpasız şirələr istisna olmaqla).
Yüksək kalorili məzmuna malik olmadan, əksər tərəvəz və meyvələr, lakin həzm olunmayan karbohidratların yüksək tərkibinə görə, tez və kifayət qədər davamlı toxluq hissinə kömək edir: pəhriz lifi çoxlu maye udmaq qabiliyyətinə malik olduğundan, onlar şişirirlər. mədə, həcminin bir hissəsini doldurun - və nəticədə doyma daha sürətli baş verir. Liflərin özləri bədənə bir kalori daşımır.
Liflərin dəyəri ondan ibarətdir ki, gündəlik qidalanmanın kifayət qədər həcmli komponenti olmaqla, insan orqanizmi tərəfindən həzm olunmur. Böyük miqdarda lifin olması yeməyin ümumi həzmini bir qədər azaldır. Lakin onun tam olmaması mədə-bağırsaq traktının işinə zərərli təsir göstərir.
Lif bağırsağın düzgün peristaltikasına (divarlarının hərəkətinə) səbəb olur və bununla da qidanın həzm kanalı vasitəsilə hərəkətinə və həzm olunmamış qidaların bədəndən çıxarılmasına kömək edir.
Qidada lazımi miqdarda lif gündəlik qida rasionunda heyvan və bitki məhsullarının düzgün kombinasiyası ilə təmin edilir.
Parçalandıqdan sonra lif, digər polisaxaridlər kimi şəkərə çevrilir. Ancaq insanın həzm sistemində belə bir parçalanmanı həyata keçirə biləcək fermentlər yoxdur. Bağırsaqlardakı mikroorqanizmlərin təsiri altında onun yalnız kiçik bir hissəsi həzm oluna bilər, lakin əsas hissəsi dəyişmədən bədəndən çıxarılır. Bu xarici faydasızlıq sayəsində lif və pektinlər balast maddələri adlanır.
Balast maddələri həzm prosesində də mühüm funksiyanı yerinə yetirir: liflər bağırsaq bakteriyaları tərəfindən mayalanır və sözün əsl mənasında yeməyi üyütməyə kömək edir; bağırsaq divarlarının sinir uclarını qıcıqlandıraraq peristaltikanı artırırlar. Qida balast maddələrində zəifdirsə, bağırsaq hərəkətliliyi pozulur, buna görə də bu pozğunluqların qarşısını almaq üçün liflə zəngin olan kobud yemlərdən istifadə etmək tövsiyə olunur.
Bundan əlavə, pəhriz lifi maddələr mübadiləsini stimullaşdırmaq qabiliyyətinə malikdir, çünki lif qida ilə gələn və ya emal zamanı əmələ gələn toksinlərin udulmasının qarşısını alır və bir növ çırpma rolunu oynayır: həzm sistemi boyunca hərəkət edərək, özləri ilə hər şeyi aparırlar. divarlara yapışıb bədəndən çıxarın.
Pəhriz lifinin başqa bir üstünlüyü, endogen xolesterol səviyyəsini azaltma qabiliyyətinə sahib olmasıdır (bu, qida ilə bizə daxil olmayan, ancaq bağırsaqlardan qaraciyərə daxil olan öd turşularından bədənin özü tərəfindən qaraciyərdə istehsal olunan xolesterindir. ).
Hemiselüloz: lif və ya sellüloza kimi taxıl məhsullarının hüceyrə divarlarının bir hissəsidir və az miqdarda meyvə və tərəvəzlərin pulpasında olur. Suyu saxlamağa və metalları bağlamağa qadirdir.
Yağ turşularının oksidləşməsi (beta oksidləşməsi). Rol H.S. – Ko bu prosesdə. Steorik turşunun tam oksidləşmə enerjisi CO 2 c H 2 O . Oksidləşmə zamanı əmələ gələn ATP molekullarının sayını hesablayın.
FA aktivləşməsi sitoplazmada, beta-oksidləşmə isə mitoxondriyada baş verir.
Acyl-CoA mitoxondrial membrandan keçə bilməz. Buna görə də, "karnitin" maddəsinin iştirakı ilə FA-nın sitoplazmadan mitoxondriyə daşınması üçün xüsusi bir mexanizm mövcuddur. Mitoxondriyanın daxili membranında ötürülməni təmin edən xüsusi nəqliyyat zülalı var. Bunun sayəsində asilkarnitin asanlıqla mitoxondrial membrana nüfuz edir.
Sitoplazmatik və mitoxondrial karnitin asiltransferazaları strukturuna görə fərqlidirlər və kinetik xüsusiyyətlərinə görə də bir-birindən fərqlənirlər. Sitoplazmatik asilkarnitin transferazanın Vmax-ı mitoxondrial fermentin Vmax-dan, həmçinin β-oksidləşmə fermentlərinin Vmax-dan aşağıdır. Buna görə də, sitoplazmik asilkarnitin transferaza yağ turşularının parçalanmasında əsas fermentdir.
Əgər yağ turşusu mitoxondriyaya daxil olarsa, o, mütləq asetil-KoA-ya katabolizm keçir.
Bədənin enerji ehtiyaclarını ödəyən ən yığcam "yanacaq" kimyəvi quruluşunun xüsusiyyətləri ilə müəyyən edilən yağ turşularıdır. 1 mol üçün yağ turşularının tam oksidləşməsi karbohidratların oksidləşməsindən bir neçə dəfə çox istifadə edilə bilən kimyəvi enerji buraxır; məsələn, 1 mol palmitik turşunun oksidləşməsi nəticəsində 130 mol ATP, 1 mol qlükozanın oksidləşməsi isə 38 mol ATP əmələ gətirir. Vahid çəkiyə görə enerji çıxışı da iki dəfədən çox fərqlənir (1 q yağ üçün 9 kkal, 1 q karbohidrat və ya zülal üçün 4 kkal). Bu yüksək enerji məhsuldarlığı benzin, neft və digər neft məhsullarını istilik və mexaniki enerji yaratmaq üçün belə effektiv yanacaq edən eyni səbəbə, yəni uzun alkil zəncirlərində karbonun yüksək dərəcədə reduksiyasına əsaslanır. Yağ turşusu molekulunun əsas hissəsi təkrarlanan vahidlərdən (CH2)n, yəni hidrogenlə maksimum zənginləşdirilmiş bir quruluşdan ibarətdir. Əvvəlki təqdimatdan da gördüyümüz kimi, bioloji oksidləşmə prosesləri zamanı yığılan enerji əsasən ADP-nin ATP-yə fosforlaşması ilə birlikdə tənəffüs zəncirinin hidrogen atomlarından elektronların idarə olunan ötürülməsi ilə əlaqədar formalaşır. Yağ turşuları ilk növbədə karbon və hidrogendən ibarət olduğundan və beləliklə, karbohidratlardan əhəmiyyətli dərəcədə az oksigen atomu ehtiva etdiyindən, yağ turşularının oksidləşməsi proporsional olaraq daha çox oksigenin udulması və deməli, oksidləşdirici fosforlaşma zamanı daha çox ATP əmələ gəlməsi ilə müşayiət olunur.
Müəyyən edilmişdir ki, yağ turşularının oksidləşməsi ən intensiv şəkildə qaraciyərdə, böyrəklərdə, skelet və ürək əzələlərində, piy toxumasında baş verir. Beyin toxumasında yağ turşularının oksidləşmə sürəti çox aşağıdır, çünki Beyin toxumasında əsas enerji mənbəyi qlükozadır.
β-oksidləşmə yağ turşusunun katabolizminin xüsusi yoludur, burada 2 karbon atomu asetil-KoA şəklində yağ turşusunun karboksil ucundan ardıcıl olaraq ayrılır. Metabolik yol - β-oksidləşmə - belə adlandırılmışdır, çünki yağ turşularının oksidləşmə reaksiyaları β-karbon atomunda baş verir. TCA dövründə asetil-KoA-nın β-oksidləşməsi və sonrakı oksidləşməsi reaksiyaları oksidləşdirici fosforlaşma mexanizmi vasitəsilə ATP sintezi üçün əsas enerji mənbələrindən biri kimi xidmət edir. Yağ turşularının β-oksidləşməsi yalnız aerob şəraitdə baş verir.
Yağ turşularının aktivləşdirilməsi
Müxtəlif reaksiyalara girməzdən əvvəl yağ turşuları aktivləşdirilməlidir, yəni. koenzim A ilə makroergik əlaqə ilə bağlıdır:
RCOOH + HSKoA + ATP → RCO ~ CoA + AMP + PPi.
Reaksiya asil-CoA sintetaza fermenti tərəfindən katalizlənir. Reaksiya zamanı ayrılan pirofosfat pirofosfataza fermenti ilə hidroliz olunur: H 4 P 2 O 7 + H 2 O → 2 H 3 PO 4.
Pirofosfatın yüksək enerjili bağının hidrolizi zamanı enerjinin ayrılması reaksiyanın tarazlığını sağa sürüşdürür və aktivləşmə reaksiyasının tamlığını təmin edir.
Asil-KoA sintetaza həm sitozolda, həm də mitoxondrial matriksdə olur. Bu fermentlər müxtəlif karbohidrogen zəncir uzunluqlarına malik yağ turşuları üçün spesifikliyi ilə fərqlənir. Qısa və orta zəncir uzunluğuna malik (4-dən 12 karbon atomuna qədər) yağ turşuları diffuziya yolu ilə mitoxondrial matrisə nüfuz edə bilir. Bu yağ turşularının aktivləşməsi mitoxondrial matrisdə baş verir. İnsan orqanizmində üstünlük təşkil edən uzun zəncirli yağ turşuları (12-20 karbon atomu) mitoxondriyanın xarici membranında yerləşən asil-CoA sintetazaları tərəfindən aktivləşdirilir.
Aktivləşdirilmiş yağ turşularının parçalanması fərziyyəyə uyğun olaraq baş verir b - oksidləşmə F. Knoop, 1904-cü ildə təklif edilmişdir b - oksidləşmə mitoxondriyada baş verir
β- Yağ turşularının oksidləşməsi- mitoxondrial matrisdə yalnız aerob şəraitdə baş verən və asetil-KoA-nın əmələ gəlməsi ilə bitən yağ turşusu katabolizminin spesifik yolu. β-oksidləşmə reaksiyalarından hidrogen CPE-yə daxil olur və asetil-KoA sitrat dövrəsində oksidləşir ki, bu da CPE-ni hidrogenlə təmin edir. Buna görə də, yağ turşularının β-oksidləşməsi tənəffüs zəncirində ATP sintezini təmin edən ən vacib metabolik yoldur.
β-oksidləşmə asil-CoA-nın FAD-dan asılı olan asil-CoA dehidrogenaz tərəfindən dehidrogenləşməsi ilə başlayır, reaksiya məhsulunda, enol-CoA-da α və β karbon atomları arasında ikiqat bağ əmələ gətirir. Bu reaksiyada bərpa olunan FADH 2 koenzimi CPE-dəki hidrogen atomlarını Q koenziminə köçürür. Nəticədə 2 ATP molekulu sintez olunur (şək. 8-27). Aşağıdakı p-oksidləşmə reaksiyasında ikiqat əlaqənin yerində su molekulu əlavə edilir ki, OH qrupu asilin β-karbon atomunda yerləşərək β-hidroksiasil-CoA əmələ gətirir. β-hidroksiasil-CoA daha sonra NAD+-dan asılı dehidrogenaz tərəfindən oksidləşir. CPE-də oksidləşən azaldılmış NADH, 3 ATP molekulunun sintezi üçün enerji təmin edir. Nəticədə β-ketoasil-KoA tiolaza fermenti tərəfindən tiolitik parçalanmaya məruz qalır, çünki C-C bağının parçalanma yerində kükürd atomu vasitəsilə koenzim A molekulu əlavə olunur.Bu 4 reaksiya ardıcıllığı nəticəsində a. iki karbonlu qalıq, asetil-CoA, asil-CoA-dan ayrılır. 2 karbon atomu ilə qısaldılmış yağ turşusu yenidən dehidrogenləşmə, nəmlənmə, dehidrogenləşmə və asetil-KoA-nın aradan qaldırılması reaksiyalarına məruz qalır. Bu reaksiyalar ardıcıllığı adətən "β-oksidləşmə dövrü" adlanır, yəni eyni reaksiyalar bütün turşu asetil qalıqlarına çevrilənə qədər yağ turşusu radikalı ilə təkrarlanır.
β -Yağ turşularının oksidləşməsi.
b-oksidləşmə prosesi siklikdir. Dövrün hər bir inqilabı üçün 2 karbon atomu asetil qalığı şəklində yağ turşusundan ayrılır.
Bundan sonra, 2 karbon atomu ilə qısaldılmış asil-CoA yenidən oksidləşməyə məruz qalır (b-oksidləşmə reaksiyalarının yeni dövrünə daxil olur). Yaranan Asetil-KoA daha sonra trikarboksilik turşu dövrünə daxil ola bilər.Siz yağ turşularının parçalanmasından əldə edilən enerji məhsuldarlığını hesablamağı bacarmalısınız. Təqdim olunan düstur n karbon atomu olan istənilən doymuş yağ turşusu üçün doğrudur.Doymamış yağ turşularının parçalanması daha az ATP istehsal edir. Yağ turşusunda hər ikiqat bağ 2 ATP molekulunun itirilməsi deməkdir. b-oksidləşməsi ən çox əzələ toxumasında, böyrəklərdə və qaraciyərdə baş verir. FA-nın b-oksidləşməsi nəticəsində Asetil-KoA əmələ gəlir. Oksidləşmə sürəti lipoliz proseslərinin sürəti ilə müəyyən edilir. Lipolizin sürətlənməsi karbohidrat aclığı və intensiv əzələ işi üçün xarakterikdir. B-oksidləşmənin sürətlənməsi qaraciyər də daxil olmaqla bir çox toxumalarda müşahidə olunur. Qaraciyər lazım olduğundan daha çox Asetil-KoA istehsal edir. Qaraciyər “altruist orqan”dır və buna görə də qaraciyər qlükozanı digər toxumalara göndərir.
Qaraciyər öz Asetil-KoA-nı digər toxumalara göndərməyə çalışır, lakin bunu edə bilmir, çünki hüceyrə membranları Asetil-KoA-nı keçirməzdir. Buna görə də qaraciyərdə Asetil-KoA-dan “keton cisimləri” adlanan xüsusi maddələr sintez olunur. Keton cisimləri asetil-KoA-nın xüsusi nəqliyyat formasıdır.
Yağ turşusu molekulu asetil koenzim A (asetil-KoA) şəklində iki karbonlu fraqmentlərin tədricən aradan qaldırılması ilə mitoxondrilərə parçalanır.
C17H35COOH + 26 O2 = 18 CO2 + 18 H2O.
Stearin turşusu oksidləşdikdə hüceyrə 146 ATP molekulu alacaq.
Yağ turşularının oksidləşməsi qaraciyərdə, böyrəklərdə, skelet və ürək əzələlərində, piy toxumasında baş verir.
F.Knoop bədən toxumalarında yağ turşusu molekulunun oksidləşməsinin b-oksidləşmədə baş verdiyini irəli sürdü. Nəticədə, karboksil qrupundan olan iki karbonlu fraqmentlər yağ turşusu molekulundan ayrılır. Yağ turşularının b-oksidləşməsi prosesi aşağıdakı mərhələlərdən ibarətdir:
Yağ turşularının aktivləşdirilməsi.Şəkər qlikolizinin birinci mərhələsinə bənzər olaraq, yağ turşuları b-oksidləşmədən əvvəl aktivləşməyə məruz qalır. Bu reaksiya ATP, koenzim A (HS-CoA) və Mg 2+ ionlarının iştirakı ilə mitoxondrial membranın xarici səthində baş verir. Reaksiya asil-CoA sintetaza tərəfindən kataliz edilir:
Reaksiya nəticəsində yağ turşusunun aktiv forması olan asil-CoA əmələ gəlir.
Yağ turşularının mitoxondriyaya daşınması. Yağ turşusunun koenzim forması, eləcə də sərbəst yağ turşuları, əslində onların oksidləşməsinin baş verdiyi mitoxondriyaya nüfuz etmək qabiliyyətinə malik deyil; karnitin (g-trimetilamino-b-hidroksibutirat) daşıyıcı kimi xidmət edir. daxili mitoxondrial membran vasitəsilə aktivləşdirilmiş yağ turşuları):
Asilkarnitin mitoxondrial membrandan keçdikdən sonra əks reaksiya baş verir - HS-CoA və mitoxondrial karnitin asiltransferazanın iştirakı ilə asilkarnitinin parçalanması:
Mitoxondriyadakı Acyl-CoA b-oksidləşmə prosesindən keçir.
Bu oksidləşmə yolu b mövqeyində yerləşən yağ turşusunun karbon atomuna oksigen atomunun əlavə edilməsini əhatə edir:
b-oksidləşmə zamanı yağ turşusunun karbon zəncirinin karboksil ucundan asetil-KoA şəklində iki karbonlu fraqmentlərin ardıcıl olaraq xaric edilməsi və müvafiq olaraq yağ turşusu zəncirinin qısaldılması baş verir:
Mitoxondrial matrisdə asil-KoA dörd reaksiyanın təkrarlanan ardıcıllığı nəticəsində parçalanır (şək. 8).
1) asil-CoA dehidrogenazın (FAD-dan asılı dehidrogenaz) iştirakı ilə oksidləşmə;
2) enol-CoA hidratazası ilə katalizləşən hidratasiya;
3) 3-hidroksiasetil-KoA dehidrogenazın (NAD-dən asılı dehidrogenaz) təsiri altında ikinci oksidləşmə;
4) asetil-CoA asiltransferazanın iştirakı ilə tioliz.
Bu dörd reaksiya ardıcıllığının cəmi yağ turşusu b-oksidləşməsinin bir dövriyyəsini təşkil edir (bax. Şəkil 8).
Nəticədə yaranan asetil-CoA Krebs dövründə oksidləşməyə məruz qalır və iki karbon atomu ilə qısaldılmış asetil-CoA, sonda butiril-CoA (4-karbonlu birləşmə) əmələ gələnə qədər yenidən bütün b-oksidləşmə yolundan keçir. b-oksidləşmə mərhələsində asetil-KoA-nın iki molekuluna parçalanır.
Tərkibində n karbon atomu olan yağ turşusu oksidləşdikdə b-oksidləşmənin n/2-1 dövrü baş verir (yəni, butiril-KoA-nın oksidləşməsi dərhal iki molekul asetil-CoA əmələ gətirdiyi üçün n/2-dən bir dövr az) və cəmi n/2 asetil-KoA molekulu alınacaq.
Məsələn, palmitik turşunun (C 16) oksidləşməsi zamanı 16/2-1 = 7 b-oksidləşmə dövrü təkrarlanır və 16/2 = 8 asetil-KoA molekulu əmələ gəlir.
Şəkil 8 – Yağ turşusunun b-oksidləşməsinin sxemi
Enerji balansı. b-oksidləşmənin hər dövrü ilə bir FADH 2 molekulu (bax. Şəkil 8; reaksiya 1) və bir NADH + H + molekulu (reaksiya 3) əmələ gəlir. Sonuncu, tənəffüs zəncirinin oksidləşməsi və əlaqəli fosforlaşma prosesində verir: FADH 2 - 2 ATP molekulu və NADH + H + - 3 ATP molekulu, yəni. bir dövrədə cəmi 5 ATP molekulu əmələ gəlir. Palmitik turşunun oksidləşməsi nəticəsində 5*7=35 ATP molekulu əmələ gəlir. Palmitik turşunun b-oksidləşməsi prosesində 8 asetil-KoA molekulu əmələ gəlir, bunların hər biri Krebs dövründə "yanaraq" 12 ATP molekulu, 8 molekul isə 12 * 8 = 96 ATP molekulu istehsal edəcəkdir.
Beləliklə, palmitik turşunun tam b-oksidləşməsi ilə ümumilikdə 35 + 96 = 131 ATP molekulu əmələ gəlir. Yağ turşusunun aktivləşdirilməsi mərhələsində ən başlanğıcda sərf olunan bir ATP molekulunu nəzərə alsaq, bir palmitik turşu molekulunun tam oksidləşməsi üçün ümumi enerji məhsuldarlığı 131-1 = 130 ATP molekulu olacaqdır.
Bununla belə, yağ turşularının b-oksidləşməsi nəticəsində əmələ gələn asetil-KoA nəinki Krebs dövrünə daxil olaraq CO 2, H 2 O, ATP-yə oksidləşə bilər, həm də xolesterolun sintezi üçün istifadə edilə bilər. qlioksilat dövründə karbohidratlar kimi.
Glyoxylate yolu yalnız bitki və bakteriyalara xasdır, heyvan orqanizmlərində yoxdur. Yağlardan karbohidratların sintezinin bu prosesi “Karbohidratların, yağların və zülalların mübadilə proseslərinin qarşılıqlı əlaqəsi” metodik göstərişində ətraflı təsvir edilmişdir (bax bənd 2.1.1, səh. 26).
Nazik bağırsağın epitel hüceyrələrindən xilomikronlar şəklində olan trigliseridlər qaraciyərə, ağciyərlərə, ürəyə, əzələlərə və digər orqanlara daxil olur, burada qliserin və yağ turşularına hidroliz olunur. Sonuncu kimi tanınan yüksək ekzerqonik metabolik yolda oksidləşə bilər